Como um estudo participativo com fruticultores exportadores, pesquisadores e comerciantes do Brasil, Chile e Reino Unido desenvolveu inovações agroecológicas para aumentar a sustentabilidade e reduzir o impacto sobre a biodiversidade.

A intensificação e a expansão agrícola são os principais fatores relacionados à crescente perda global de biodiversidade observada neste século. Esse fenômeno é relevante e impacta países do sul global, incluindo os da América do Sul, que se destacam na produção agropecuária e exportação de commodities. Neste contexto, o plano Global da Biodiversidade apoia a adoção de inovações agroecológicas para reduzir os impactos da agricultura sobre a biodiversidade. As inovações agroecológicas são práticas recentemente introduzidas nos sistemas de manejo agrícola que possuem o potencial de favorecer, em paisagens agrícolas e a longo prazo, a biodiversidade e os benefícios que ela promove em áreas cultivadas.
Para que tais esforços sejam eficazes é preciso que os agricultores estejam preparados e motivados a desenvolver inovações agroecológicas que se adequem a sistemas produtivos e eficientes, que ajudem a minimizar o uso de terras para a produção de alimentos. Também é preciso que os agricultores estejam engajados e empoderados para adotar tais inovações, de modo a vislumbrar oportunidades diante do cenário político global e de alguns incentivos de mercados internacionais, a exemplo do mercado europeu. No entanto, as ações de cima para baixo que incentivam a sustentabilidade são insuficientes, ou sucumbem diante do contexto sistêmico que favorece modelos de produção intensivos.
De modo a entender como promover a participação ativa de agricultores convencionais de grande e médio porte, o projeto Manejo Sustentável de Fruteiras na Caatinga – SUFICA (www.sufica.org), utilizou a abordagem transdisciplinar e participativa descrita em estudo publicado recentemente. O nosso modelo de coprodução de conhecimento (Figura 1) ilustra como a investigação transdisciplinar pode permitir aos agricultores do Sul global envolverem-se na conservação da biodiversidade e produção sustentável em fazendas de fruticultura.
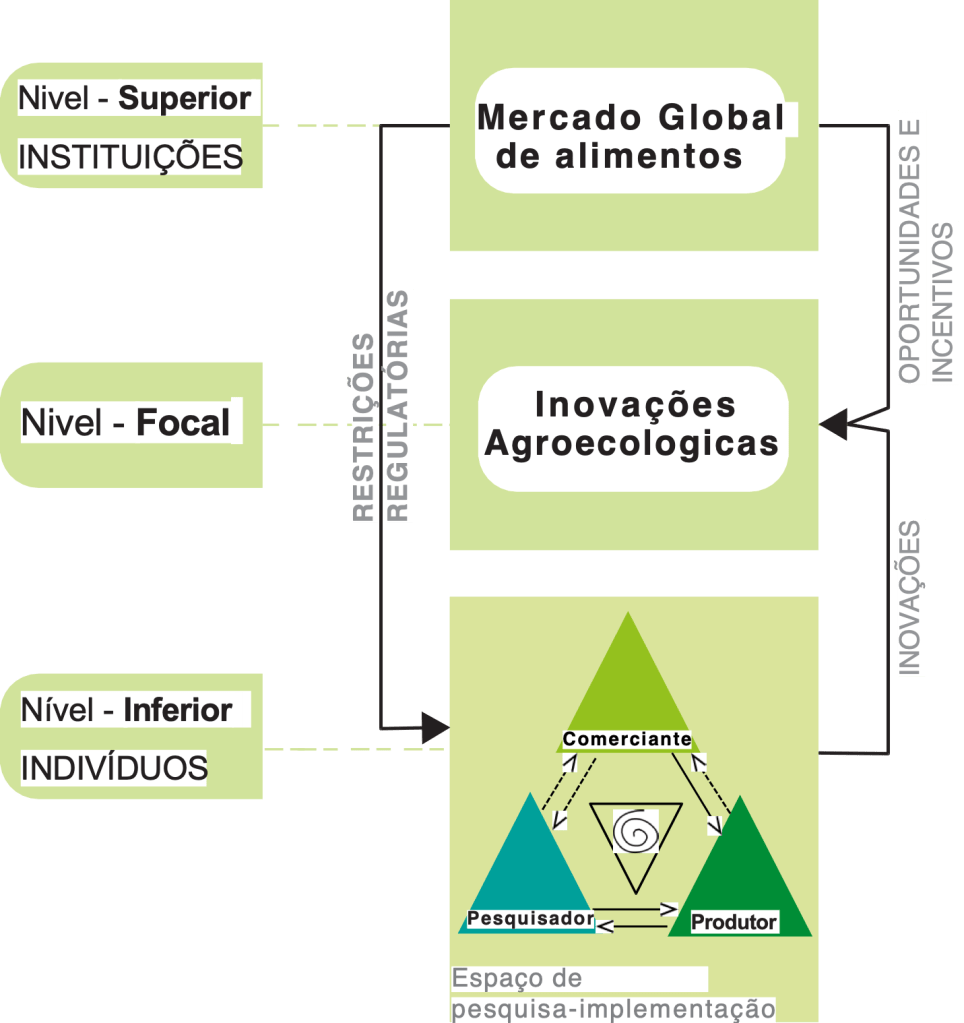
Figura 1: Modelo hierárquico em três níveis ilustrando a relação entre o mercado global de alimentos e a emergência de inovações agroecológicas. Essas inovações são testadas pelos agricultores, sob as regulamentações, restrições, oportunidades e incentivos do mercado global de alimentos (nível superior). O processo participativo, envolvendo agricultores, representantes da indústria de alimentos, e pesquisadores, exercem papel crucial na tomada de decisão sobre a adoção das práticas agroecológicas na agricultura (nível focal). Dentro do “espaço iterativo” (nível inferior) ocorre a coprodução de conhecimento pelos agricultores, representantes da industria de alimentos e pesquisadores, conduzindo ao desenvolvimento, implementação e usabilidade das inovações agroecológicas. Este processo de baixo para cima (bottom-up mechanism) promove a emergência de inovações agroecológicas com base cientifica e de fácil implementação (Adaptado de Salthe, 2010, 2012; Rocha & Rocha, 2018). Imagem desenhada por Germana Gonçalves de Araujo. Fonte: https:// doi.org/10.1002/pan3.10613
Durante o estudo, desenvolvido durante quatro anos, contamos com a participação de 14 agricultores, abrangendo uma área significativa ocupada por fruticultura intensiva (por exemplo, uva de mesa, manga e cereja) voltada, principalmente, para a exportação. Todos os agricultores estavam pressionados por compradores internacionais, especialmente da União Européia, a apresentarem ações voltadas à conservação da biodiversidade. Nosso estudo se concentrou em áreas com elevada biodiversidade, afetadas pela perda de habitat, situadas em região mediterrânea no Chile e de floresta tropical seca (Caatinga) no Brasil.
Os agricultores colaboraram ativamente com os pesquisadores e representantes da indústria, em um processo iterativo de diálogo e grupo de trabalho. Os agricultores participaram de todas as etapas do estudo, com diferentes niveis de participação, desde consultivo até a coprodução das inovações agroecológicas. Após avaliarem as evidências da Conservation Evidence (2023)1 e a experiência local, três inovações agroecológicas foram escolhidas: cultivo de cobertura, faixas com plantas nativas, e poleiros para aves. Estas inovações, priorizadas pelos produtores entre outras opções listadas (Figura 2), foram implementadas por nove produtores (apesar da Covid!).

Figura 2: Inovações agroecológicas implementadas nas fazendas. Poleiros para aves usado pela espécie alvo: (a) “gavião” (Parabuteo unicinctus) fotografado com armadilha de camera no Chile; (b) “carcará” (Caracara plancus) fotografado por um produtor no Brasil; (c) cultivo de cobertura entre as linhas do vinhedo no Brasil e (d) faixa de planta nativa no Chile. Créditos da fotos: (a,d) Nadia Rojas- Arévalo, (b) agricultor participante anônimo, (c) Patricia Oliveira-Rebouças. Fonte: https://doi.org/10.1002/pan3.10613
Nós conduzimos uma série de atividades ao longo de quatro anos, tais como oficinas participativas presenciais, em grupo e individuais, e também realizamos conversas via online, para manter a comunicação e compartilhar boas práticas, visando superar os desafios impostos pela distância geográfica e diversidade de participantes. Por meio dessas atividades conseguimos aumentar progressivamente o nível de participação, de modo que alguns agricultores engajados realizaram o monitoramento ativo da efetividade das práticas e sugeriram adaptações adequadas às suas condições particulares. As inovações agroecológicas implementadas foram mantidas após a finalização do projeto, pois ao menos uma delas foi adotada por oito dentre nove fazendas. Os agricultores relataram que a principal motivação para manter as práticas foi seu alinhamento com o sistema de manejo das fazendas e por poderem ser reconhecidas como atitude positiva à conservação da biodiversidade em sua cadeia produtiva. Além das práticas testadas neste estudo, o processo participativo disponibilizou uma lista de práticas agroecológicas, selecionada com base em evidência científica, adequação ao contexto e com boa relação custo-benefício (Conservation Evidence, 2023) que podem ser testadas em fazendas de fruticultura de regiões semiáridas de países da América do Sul, visando impulsionar a transição para um modelo de agricultura amigável à biodiversidade na região.
Ao longo da pesquisa, desenvolvemos juntos um conjunto amplo de recursos para a disseminação do conhecimento gerado a tomadores de decisão e compradores influentes localmente, os quais podem atuar como facilitadores do conhecimento e ampliar o alcance do conhecimento gerado localmente. Estes recursos incluem uma métrica de sustentabilidade online, uma série com seis cartilhas, que estão disponíveis, acessíveis e traduzidas para o idioma dos participantes, e vídeos (por exemplo, https://zenodo.org/records/10070493 ).
A nossa experiência com a realização deste projeto participativo com os agricultores evidenciou que as inovações agroecológicas têm mais chances de serem usadas quando selecionadas pelos agricultores. A partir dos resultados deste estudo, destacamos a importância de abordagens de pesquisa transdisciplinar que enfatizam a coprodução de conhecimento de base local e a colaboração entre várias partes interessadas. Sugerimos que esforços de baixo para cima para impulsionar a sustentabilidade devem ser priorizados em relação aos esforços de cima para baixo. Defendemos que as abordagens de base local podem beneficiar estratégias de cima para baixo, tais como incentivos de mercado, códigos voluntários ou regulamentações comerciais na agricultura, contribuindo para alcançar maior eficácia e promover a transição agroecológica2. Também destacamos que as partes interessadas, ligadas à produção agrícola e à indústria alimentar, podem se beneficiar de trabalhos em parceria com pesquisadores locais.
Fabiana Oliveira da Silva
Departamento de Educação em Ciências Agrárias e da Terra, Universidade Federal de Sergipe – UFS
Eduardo C. Arellano
Facultad de Agronomía y Sistemas Naturales e Instituto para el Desarrollo Sustentable, Pontificia Universidad Católica de Chile/ Center of Applied Ecology and Sustainability – CAPES
Blandina Felipe Viana
Universidade Federal da Bahia – UFBA
Vini Gbami Silva Ferreira
Universidade Federal do Vale do São Francisco – UNIVASF
Patricia L. Oliveira Rebouças
Departamento de Tecnologia e Ciências Sociais, Universidade do Estado da Bahia – UNEB Campus III
Nadia Rojas-Arévalo
Department of Earth and Environmental Sciences, Faculty of Science and Engineering, University of Manchester
Andrés Muñoz- Sáez
Departamento de Producción Agrícola, Facultad de Ciencias Agronómicas, Universidad de Chile
Valentina P. Jimenez
Center of Applied Ecology and Sustainability
Natalia B. Zielonka
Durrell Institute of Conservation and Ecology, University of Kent and School of Biological Sciences, University of East Anglia
Liam P. Crowther
School of Biological Sciences, University of East Anglia)
Lynn V. Dicks
Department of Zoology, Conservation Research Institute, University of Cambridge
PARA SABER MAIS
Oliveira da Silva, F., Arellano, E. C., Felipe Viana, B., Silva-Ferreira, V., Oliveira-Rebouças, P., Rojas-Arévalo, N., Muñoz-Sáez, A., Jimenez, V. P., Zielonka, N. B., Crowther, L. P., & Dicks, L. V. (2024). Co-production of agroecological innovations to improve sustainability in South American fruit farms. People and Nature, 00, 1–16. https://doi.org/10.1002/pan3.10613
NOTAS
- Conservation Evidence (www.conservationevidence.com) é um recurso disponível online, gratuito e confiável, que permite o acesso fácil a ainformações mais recentes e relevantes , criado para apoiar decisões sobre como manter e restaurar a biodiversidade global. Fornece evidências e uma avaliação dessas evidências, que devem ser interpretadas por conservacionistas que compreendam seu próprio local e a situação nacional ou regional. ↩︎
- Processo em que sistemas produtivos convencionais da agricultura moderna, passam gradualmente a adotar princípios e tecnologias de base ecológica, tornando-se agroecossistemas mais sustentáveis. (https://www.atermaisdigital.cnptia.embrapa.br/web/saf/transicao-agroecologica)%5D. ↩︎