Muitas aves se reproduzem em um local e, durante o outono e inverno, percorrem longas distâncias em busca de alimento em regiões onde as condições são mais favoráveis. Conheça um pouco mais sobre esse fenômeno fascinante, e como seu estudo pode conectar cientistas e cidadãos em ações de conservação e produção de conhecimento.
A vida na Terra é regida por ciclos, dos nutrientes que circulam pelos ecossistemas, das alternâncias entre dia e noite, e das estações que se repetem anualmente. Esses ciclos funcionam como um calendário natural para diversas espécies que realizam movimentos sazonais, ou seja, deslocamentos que ocorrem em períodos específicos do ano e se repetem regularmente. Na biologia, chamamos esses movimentos de “migração”. Eles são realizados por espécies que se reproduzem em um local e, durante o outono e inverno, percorrem longas distâncias em busca de alimento em regiões onde as condições são mais favoráveis à sobrevivência.
Esses movimentos sempre fascinaram sociedades humanas, que observavam a chegada e partida das aves como sinais de mudanças no clima e, muitas vezes, como marcadores do calendário agrícola. Para os Kawaiweté, povo indígena do Xingu, a chegada das andorinhas anuncia o início da temporada de chuvas. Com o avanço do desmatamento e a consequente alteração no regime de chuvas, a presença e o número dessas aves avistadas vem mudando, revelando como mudanças ambientais afetam diretamente as migrações. Afinal, para onde as aves vão e por que partem são perguntas tão antigas quanto a própria relação humana com a natureza.
Na Grécia antiga, Aristóteles propôs uma teoria curiosa: acreditava que algumas aves desaparecidas no inverno se transformavam em outras espécies. Já durante a Idade Média, imaginava-se que andorinhas hibernavam no fundo de lagos congelados. O mistério só começou a ser desvendado séculos depois e, de certa forma, junto com outra lenda famosa: a de que cegonhas traziam bebês.
A crença estava relacionada à migração da cegonha-branca (Ciconia ciconia). Na Idade Média europeia, muitos casamentos aconteciam no início do verão. Na sequência, no início do outono, as cegonhas desapareciam e retornavam aproximadamente nove meses depois da temporada de casamentos, coincidindo com o nascimento de muitas crianças. A associação se popularizou por gerações. Até que, em 1822, algo incomum apareceu em um telhado da pequena cidade de Klütz, na Alemanha: uma cegonha com uma flecha atravessando o pescoço. Análises da origem da madeira da flecha confirmaram que ela havia viajado pelo menos 2.000 km entre a Europa e a África, e assim a compreensão moderna sobre migração ganhou um marco histórico.
A Cegonha flechada de Klütz, hoje preservada na Universidade de Rostock. A palavra em alemão Pfeilstorch (“cegonha-flecha”) foi criada para designar estes animais que chegavam à Europa atingidos por flechas ou lanças. Desde 1822, cerca de 23 desses casos foram registrados na Alemanha. Fonte: https://en.wikipedia.org/wiki/Pfeilstorch, foto do Zoologische Sammlung der Universität Rostock.
Décadas depois, em 1890, o dinamarquês Hans Mortensen inaugurou a marcação de aves com anilhas, permitindo conhecer rotas migratórias, longevidade e conectividade populacional. O método é amplamente utilizado até hoje, embora dependa de grande esforço de captura e recaptura. A partir de meados dos anos 1980, o surgimento de dispositivos que estimam posição pela luminosidade, radiotransmissores, e, posteriormente, de rastreadores GPS cada vez menores trouxe uma revolução aos estudos de migração, permitindo acompanhar trajetos completos inclusive de aves pequenas, como as andorinhas.
Existem cerca de 88 espécies de andorinhas no mundo, 18 delas no Brasil, todas apresentando algum tipo de movimento migratório. Seis espécies se reproduzem no Hemisfério Norte e chegam à América do Sul durante a nossa primavera e verão. No Brasil, andorinhas fazem parte do imaginário popular e aparecem frequentemente em praças e fiações elétricas, formando bandos expressivos e revoadas marcantes que, junto com as suas chegadas e partidas, inspiraram versos como os do Trio Parada Dura: “Nós somos andorinhas que vão e que vêm… Uma andorinha voando sozinha não faz verão”.
Bando de andorinhas-azuis (Progne subis) em Linhares, ES, Brasil. Foto: Gabriel Bonfá.
Há sete anos estudo uma dessas espécies, a andorinha-azul (Progne subis), observando os bandos que chegam ao Brasil entre setembro e abril. Conversando com moradores, percebo sempre dois pontos: encantamento com as revoadas ao amanhecer e entardecer, mas também incômodo com o barulho e o acúmulo de fezes nas áreas onde dormem; e a dúvida comum: “De onde elas vêm?”. A andorinha-azul se reproduz no sul do Canadá, nos Estados Unidos e no norte do México, mas passa a estação não reprodutiva praticamente em todo o território brasileiro. Um dos objetivos centrais da minha pesquisa é conectar cada região do Brasil às populações do Hemisfério Norte, muitas vezes separadas por dezenas de milhares de quilômetros.
Para compreender esses movimentos, é essencial conhecer a biologia da espécie. As andorinhas-azuis são divididas em três subespécies, isoladas geográfica e reprodutivamente, que diferem na distribuição, no tipo de ninho utilizado e em aspectos morfológicos sutis:
A subespécie do leste, a mais estudada, usa ninhos artificiais em jardins da metade leste dos EUA e Canadá. A facilidade de captura e recaptura permitiu rastrear centenas de indivíduos via GPS, revelando que passam o período não reprodutivo principalmente na Amazônia, onde formam imensos dormitórios com até 500 mil aves.
A subespécie do oeste nidifica em cavidades de árvores ao longo da costa oeste, do Canadá à Califórnia. Poucas populações utilizam ninhos artificiais e, por isso, poucas aves foram rastreadas, mas sabemos que aparecem durante a migração no litoral do Rio de Janeiro, Espírito Santo e Bahia.
A subespécie do deserto utiliza ocos de cactos no deserto de Sonora (Arizona, Califórnia e noroeste do México). É a menos conhecida: seu destino migratório permaneceu um mistério até 2023, quando um indivíduo foi rastreado até o Nordeste do Brasil.
A facilidade de recapturar os mesmos indivíduos no leste da América do Norte permitiu o uso de dispositivos de rastreamento que funcionam como discos de memória, nos quais os dados só podem ser acessados quando a ave retorna ao mesmo ninho e o dispositivo é retirado. Já nas outras subespécies, esse processo é muito mais desafiador: além de ocuparem cavidades naturais em árvores altas ou ocos de cactos no deserto, locais de difícil acesso, essas populações são menos gregárias durante a reprodução, o que reduz as chances de capturar e, principalmente, recapturar os mesmos indivíduos. No Brasil é relativamente fácil capturar andorinhas nos imensos bandos que se formam para dormir, mas é praticamente impossível reencontrar no ano seguinte o mesmo indivíduo para recuperar dispositivos. Nos últimos anos, surgiram transmissores que enviam dados em tempo real via satélite, porém, até 2021, eles ainda eram pesados demais para serem usados em aves tão pequenas.
Isso mudou quando um novo rastreador de apenas 2 g se tornou disponível e em 2025, um marco tornou essa conexão ainda mais clara: cinco andorinhas-azuis foram equipadas com transmissores via satélite em um grande dormitório de Linhares, permitindo acompanhar em tempo real suas trajetórias migratórias até o Novo México, a Califórnia e a Colúmbia Britânica. Esses avanços têm revelado rotas, gargalos ecológicos e diferenças entre subespécies, abrindo caminho para um mapa cada vez mais detalhado e preciso da presença e dos movimentos da espécie no Brasil.
Andorinha-azul (Progne subis) com rádiotransmissor no dorso. Foto: Dado Galdieri/Hilaea Media
A conservação da andorinha-azul depende diretamente da colaboração entre pesquisadores e comunidades locais. Em muitas cidades brasileiras, essas aves dormem em praças e áreas urbanas, o que gera conflitos, mas que pode levar a medidas inadequadas, como podas ou remoção de árvores usadas como dormitórios. Ao mesmo tempo, plataformas de ciência cidadã como eBird, WikiAves e observações diretas de moradores permitem localizar bandos rapidamente, registrar datas de chegada e partida e monitorar mudanças ao longo dos anos. Esses dados são fundamentais especialmente diante das mudanças climáticas, que já vêm alterando padrões de chuva e disponibilidade de insetos, afetando a sobrevivência das aves durante sua estadia no Brasil. Assim, a participação da população é essencial para transformar potenciais conflitos em oportunidades de convivência e proteção.
A migração das aves não reconhece fronteiras, e a conservação também não pode reconhecê-las. Proteger espécies que viajam milhares de quilômetros exige redes de cooperação entre pesquisadores, instituições, comunidades e governos. Projetos que conectam diferentes países e continentes são essenciais para mapear rotas completas, compartilhar dados de rastreamento, comparar ameaças regionais e alinhar ações de conservação. Cada recaptura, cada nova marcação e cada dormitório monitorado são peças de um esforço global para compreender as aves viajantes.
Clarissa Santos Mestre em ecologia pela Universidade Federal de São Carlos, aluna de doutorado em ecologia pela Universidade de São Paulo.
PARA SABER MAIS
Projeto Andorinha-azul: iniciativas de pesquisa que estudam migração, saúde das aves, uso de habitat e conectividade entre Brasil e América do Norte. andorinhaazul.org, Instagram: @andorinhazul
Como um estudo participativo com fruticultores exportadores, pesquisadores e comerciantes do Brasil, Chile e Reino Unido desenvolveu inovações agroecológicas para aumentar a sustentabilidade e reduzir o impacto sobre a biodiversidade.
A intensificação e a expansão agrícola são os principais fatores relacionados à crescente perda global de biodiversidade observada neste século. Esse fenômeno é relevante e impacta países do sul global, incluindo os da América do Sul, que se destacam na produção agropecuária e exportação de commodities. Neste contexto, o plano Global da Biodiversidade apoia a adoção de inovações agroecológicas para reduzir os impactos da agricultura sobre a biodiversidade. As inovações agroecológicas são práticas recentemente introduzidas nos sistemas de manejo agrícola que possuem o potencial de favorecer, em paisagens agrícolas e a longo prazo, a biodiversidade e os benefícios que ela promove em áreas cultivadas. Para que tais esforços sejam eficazes é preciso que os agricultores estejam preparados e motivados a desenvolver inovações agroecológicas que se adequem a sistemas produtivos e eficientes, que ajudem a minimizar o uso de terras para a produção de alimentos. Também é preciso que os agricultores estejam engajados e empoderados para adotar tais inovações, de modo a vislumbrar oportunidades diante do cenário político global e de alguns incentivos de mercados internacionais, a exemplo do mercado europeu. No entanto, as ações de cima para baixo que incentivam a sustentabilidade são insuficientes, ou sucumbem diante do contexto sistêmico que favorece modelos de produção intensivos.
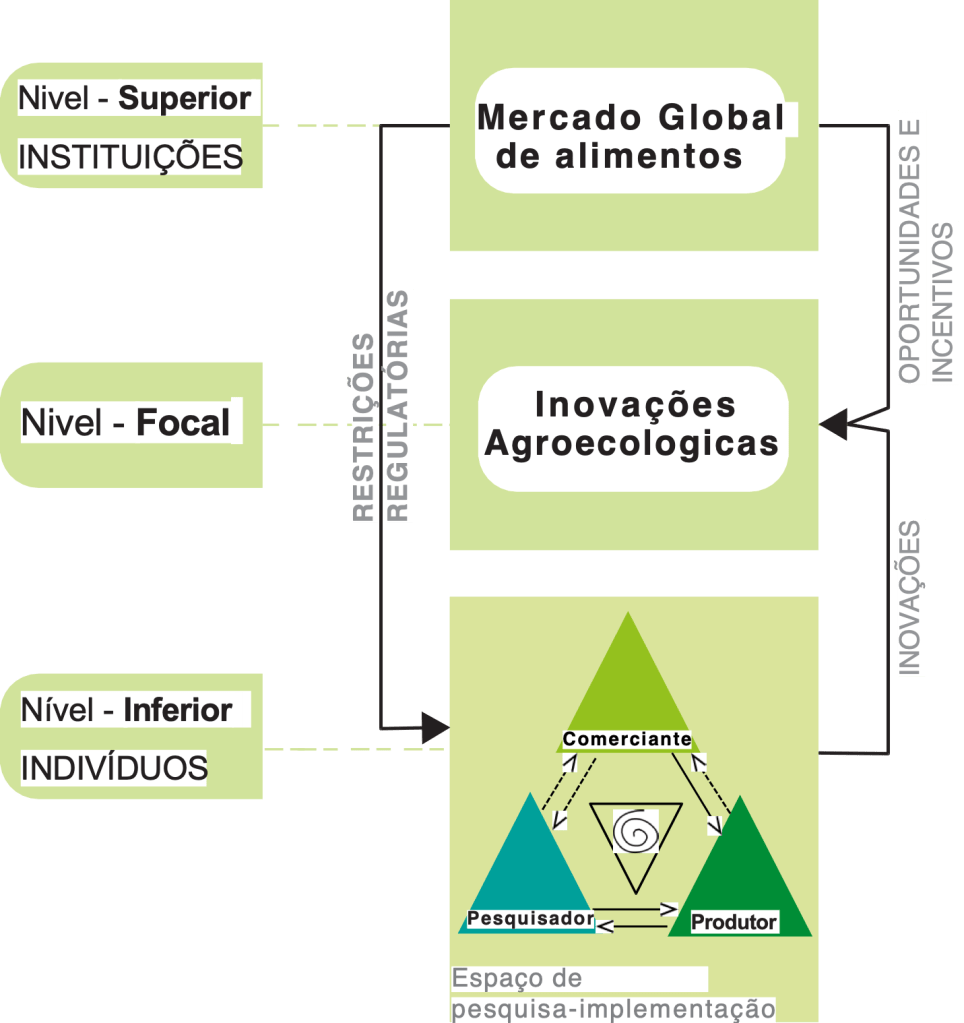
De modo a entender como promover a participação ativa de agricultores convencionais de grande e médio porte, o projeto Manejo Sustentável de Fruteiras na Caatinga – SUFICA (www.sufica.org), utilizou a abordagem transdisciplinar e participativa descrita em estudo publicado recentemente. O nosso modelo de coprodução de conhecimento (Figura 1) ilustra como a investigação transdisciplinar pode permitir aos agricultores do Sul global envolverem-se na conservação da biodiversidade e produção sustentável em fazendas de fruticultura.
Figura 1: Modelo hierárquico em três níveis ilustrando a relação entre o mercado global de alimentos e a emergência de inovações agroecológicas. Essas inovações são testadas pelos agricultores, sob as regulamentações, restrições, oportunidades e incentivos do mercado global de alimentos (nível superior). O processo participativo, envolvendo agricultores, representantes da indústria de alimentos, e pesquisadores, exercem papel crucial na tomada de decisão sobre a adoção das práticas agroecológicas na agricultura (nível focal). Dentro do “espaço iterativo” (nível inferior) ocorre a coprodução de conhecimento pelos agricultores, representantes da industria de alimentos e pesquisadores, conduzindo ao desenvolvimento, implementação e usabilidade das inovações agroecológicas. Este processo de baixo para cima (bottom-up mechanism) promove a emergência de inovações agroecológicas com base cientifica e de fácil implementação (Adaptado de Salthe, 2010, 2012; Rocha & Rocha, 2018). Imagem desenhada por Germana Gonçalves de Araujo. Fonte: https:// doi.org/10.1002/pan3.10613
Durante o estudo, desenvolvido durante quatro anos, contamos com a participação de 14 agricultores, abrangendo uma área significativa ocupada por fruticultura intensiva (por exemplo, uva de mesa, manga e cereja) voltada, principalmente, para a exportação. Todos os agricultores estavam pressionados por compradores internacionais, especialmente da União Européia, a apresentarem ações voltadas à conservação da biodiversidade. Nosso estudo se concentrou em áreas com elevada biodiversidade, afetadas pela perda de habitat, situadas em região mediterrânea no Chile e de floresta tropical seca (Caatinga) no Brasil.
Os agricultores colaboraram ativamente com os pesquisadores e representantes da indústria, em um processo iterativo de diálogo e grupo de trabalho. Os agricultores participaram de todas as etapas do estudo, com diferentes niveis de participação, desde consultivo até a coprodução das inovações agroecológicas. Após avaliarem as evidências da Conservation Evidence (2023)1 e a experiência local, três inovações agroecológicas foram escolhidas: cultivo de cobertura, faixas com plantas nativas, e poleiros para aves. Estas inovações, priorizadas pelos produtores entre outras opções listadas (Figura 2), foram implementadas por nove produtores (apesar da Covid!).
Figura 2: Inovações agroecológicas implementadas nas fazendas. Poleiros para aves usado pela espécie alvo: (a) “gavião” (Parabuteo unicinctus) fotografado com armadilha de camera no Chile; (b) “carcará” (Caracara plancus) fotografado por um produtor no Brasil; (c) cultivo de cobertura entre as linhas do vinhedo no Brasil e (d) faixa de planta nativa no Chile. Créditos da fotos: (a,d) Nadia Rojas- Arévalo, (b) agricultor participante anônimo, (c) Patricia Oliveira-Rebouças. Fonte: https://doi.org/10.1002/pan3.10613
Nós conduzimos uma série de atividades ao longo de quatro anos, tais como oficinas participativas presenciais, em grupo e individuais, e também realizamos conversas via online, para manter a comunicação e compartilhar boas práticas, visando superar os desafios impostos pela distância geográfica e diversidade de participantes. Por meio dessas atividades conseguimos aumentar progressivamente o nível de participação, de modo que alguns agricultores engajados realizaram o monitoramento ativo da efetividade das práticas e sugeriram adaptações adequadas às suas condições particulares. As inovações agroecológicas implementadas foram mantidas após a finalização do projeto, pois ao menos uma delas foi adotada por oito dentre nove fazendas. Os agricultores relataram que a principal motivação para manter as práticas foi seu alinhamento com o sistema de manejo das fazendas e por poderem ser reconhecidas como atitude positiva à conservação da biodiversidade em sua cadeia produtiva. Além das práticas testadas neste estudo, o processo participativo disponibilizou uma lista de práticas agroecológicas, selecionada com base em evidência científica, adequação ao contexto e com boa relação custo-benefício (Conservation Evidence, 2023) que podem ser testadas em fazendas de fruticultura de regiões semiáridas de países da América do Sul, visando impulsionar a transição para um modelo de agricultura amigável à biodiversidade na região.
Ao longo da pesquisa, desenvolvemos juntos um conjunto amplo de recursos para a disseminação do conhecimento gerado a tomadores de decisão e compradores influentes localmente, os quais podem atuar como facilitadores do conhecimento e ampliar o alcance do conhecimento gerado localmente. Estes recursos incluem uma métrica de sustentabilidade online, uma série com seis cartilhas, que estão disponíveis, acessíveis e traduzidas para o idioma dos participantes, e vídeos (por exemplo, https://zenodo.org/records/10070493 ).
A nossa experiência com a realização deste projeto participativo com os agricultores evidenciou que as inovações agroecológicas têm mais chances de serem usadas quando selecionadas pelos agricultores. A partir dos resultados deste estudo, destacamos a importância de abordagens de pesquisa transdisciplinar que enfatizam a coprodução de conhecimento de base local e a colaboração entre várias partes interessadas. Sugerimos que esforços de baixo para cima para impulsionar a sustentabilidade devem ser priorizados em relação aos esforços de cima para baixo. Defendemos que as abordagens de base local podem beneficiar estratégias de cima para baixo, tais como incentivos de mercado, códigos voluntários ou regulamentações comerciais na agricultura, contribuindo para alcançar maior eficácia e promover a transição agroecológica2. Também destacamos que as partes interessadas, ligadas à produção agrícola e à indústria alimentar, podem se beneficiar de trabalhos em parceria com pesquisadores locais.
Fabiana Oliveira da Silva Departamento de Educação em Ciências Agrárias e da Terra, Universidade Federal de Sergipe – UFS
Eduardo C. Arellano Facultad de Agronomía y Sistemas Naturales e Instituto para el Desarrollo Sustentable, Pontificia Universidad Católica de Chile/ Center of Applied Ecology and Sustainability – CAPES
Blandina Felipe Viana Universidade Federal da Bahia – UFBA
Vini Gbami Silva Ferreira Universidade Federal do Vale do São Francisco – UNIVASF
Patricia L. Oliveira Rebouças Departamento de Tecnologia e Ciências Sociais, Universidade do Estado da Bahia – UNEB Campus III
Nadia Rojas-Arévalo Department of Earth and Environmental Sciences, Faculty of Science and Engineering, University of Manchester
Andrés Muñoz- Sáez Departamento de Producción Agrícola, Facultad de Ciencias Agronómicas, Universidad de Chile
Valentina P. Jimenez Center of Applied Ecology and Sustainability
Natalia B. Zielonka Durrell Institute of Conservation and Ecology, University of Kent and School of Biological Sciences, University of East Anglia
Liam P. Crowther School of Biological Sciences, University of East Anglia)
Lynn V. Dicks Department of Zoology, Conservation Research Institute, University of Cambridge
PARA SABER MAIS
Oliveira da Silva, F., Arellano, E. C., Felipe Viana, B., Silva-Ferreira, V., Oliveira-Rebouças, P., Rojas-Arévalo, N., Muñoz-Sáez, A., Jimenez, V. P., Zielonka, N. B., Crowther, L. P., & Dicks, L. V. (2024). Co-production of agroecological innovations to improve sustainability in South American fruit farms. People and Nature, 00, 1–16. https://doi.org/10.1002/pan3.10613
NOTAS
Conservation Evidence (www.conservationevidence.com) é um recurso disponível online, gratuito e confiável, que permite o acesso fácil a ainformações mais recentes e relevantes , criado para apoiar decisões sobre como manter e restaurar a biodiversidade global. Fornece evidências e uma avaliação dessas evidências, que devem ser interpretadas por conservacionistas que compreendam seu próprio local e a situação nacional ou regional. ↩︎
Construir um novo perfil profissional é criar o próprio espaço no mundo
Vincent van Gogh (1853 – 1890), Paris, Dezembro de 1887-Fevereiro de 1888. Óleo sobre tela, 65.1 cm x 50 cm. Créditos: Van Gogh Museum, Amsterdam (Vincent van Gogh Foundation). O artista tinha o costume de pintar autorretratos, pois não tinha recursos para pagar modelos. Os autorretratos eram feitos para estudar técnicas e, por isso mesmo, ele aparece de diferentes maneiras nos quadros. A obra foi escolhida para ilustrar a ideia de que uma pintura de si mesmo é sempre uma releitura e, para esta reflexão, simboliza a criação desse novo perfil profissional. Além disso, Van Gogh foi responsável por criar um novo estilo artístico, inexistente à época, o que também associo a uma nova forma de fazer ciência.
Um dos maiores desafios na Ecologia e Conservação é a lacuna pesquisa-prática. Este desafio está relacionado à constatação de que o conhecimento produzido na Ecologia, bem como a forma de pensar e investigar questões ambientais, não estão sendo aplicadas para resolver problemas do mundo real. Adicionalmente a isso, na Biologia da Conservação cresce a ideia da importância de considerar a relação humano-natureza, em vez de ver os humanos como separados da natureza. No entanto, a superação dos problemas ambientais utilizando do conhecimento científico e a proposição de soluções criativas baseada em diversos saberes é um trabalho bastante difícil. Isso depende de tradução e fomento ao intercâmbio entre os conhecimentos, de envolver diferentes partes interessadas, de reunir e sistematizar conhecimento(s) de boa qualidade, de compreender novos desafios e manejar relações interpessoais.
A pesquisa inter- e transdisciplinar desempenha um papel importante nesse aspecto, configurando espaços onde diferentes perspectivas possam trazer suas narrativas sobre os problemas socioambientais, ao passo que desenvolve e reproduz práticas diferentes da ciência dominante. Criam-se, assim, soluções que potencialmente são mais benéficas para as pessoas e natureza. No entanto, quem fará todo o trabalho árduo de integrar as abordagens reunidas em iniciativas dessa natureza? Quais são as reais necessidades de quem se arrisca a iniciar uma jornada inter- e transdisciplinar? Aqui eu vou compartilhar brevemente minha própria experiência como bióloga em início de carreira que migrou de uma formação disciplinar para uma atuação inter- e transdisciplinar. Vou discutir alguns dos desafios e benefícios que encontrei e argumentar sobre porque a inter- e transdisciplinaridade são cruciais para abordar questões socioambientais.
Vamos começar do início. Durante o período de graduação, estudei Ciências Biológicas por quatro anos. Esse período foi suficiente para moldar minha forma de pensar, meu comportamento e minha visão de mundo. Eu esqueci como costumava pensar quando comecei meus estudos e achava difícil compreender porque alguém discordaria da minha perspectiva sobre a natureza – um assunto implicitamente estudado em minha área e profundamente enraizado em meu conhecimento adquirido através de incontáveis horas de estudo. Isso aconteceu, em parte, porque minha formação acadêmica tinha um foco disciplinar muito forte. Aprendi a evolução como um conceito central na Biologia e subjacente à diversidade, taxonomia, genética, zoologia, botânica e ecologia. Durante esse tempo de graduação, cada disciplina era dividida em dois ou três módulos semestrais, mais ou menos conectados entre si. Eu passei da bioquímica e biologia molecular para a genética e, paralelamente, estudei embriologia, zoologia, botânica e ecologia. Cada uma em suas caixas e com sua relevância. No final da minha graduação, eu tinha um diploma que certificava meu conhecimento em várias disciplinas dentro do campo da biologia. No entanto, embora os seres humanos fossem considerados parte da natureza, o aspecto “humano” não foi abordado de forma abrangente.
Da metade para o final da minha graduação, eu li o texto intitulado “Desafios e Oportunidades de Superar a Lacuna entre a Pesquisa e a Implementação na Ciência Ecológica e Gestão no Brasil”, escrito por pessoas que eu admiro muito por seu excelente trabalho em Ecologia e Filosofia da Ciência. Ainda nesse período, tive a sorte de fazer parte de um grande projeto de pesquisa em que a lacuna entre pesquisa e implementação era foco de discussões profundas e de vários esforços para tornar a Ecologia mais eficaz na resolução de problemas do mundo real, notadamente na agricultura, mas também com comunidades pesqueiras, manejo de áreas preservadas e políticas públicas. Este foi um momento de virada, quando percebi que as questões ambientais com as quais eu me preocupava tinham facetas que não poderiam ser adequadamente estudadas dentro da minha formação disciplinar, devido às limitações metodológicas, falta de ferramentas ou paradigmas prevalentes. De fato, as universidades são tipicamente instituições orientadas por disciplinas e apenas recentemente a área de Conservação começou a abraçar uma perspectiva de “natureza e pessoas” de forma mais ampla, o que tem implicações para a gestão, para a avaliação de currículo e impactos científicos, bem como para o desenvolvimento de teorias, métodos e ferramentas que sirvam para compreender e avaliar os sistemas socioecológicos.
Para enfrentar esse problema, percebi que precisava entender outras abordagens fora da minha formação acadêmica e começar uma jornada inter- e transdisciplinar. Antes de aprofundar ainda mais essa discussão, é muito importante esclarecer as diferenças entre estudos disciplinares, multidisciplinares, interdisciplinares e transdisciplinares. Aqui, vou adotar os conceitos apresentados no artigo de Tress, Tress e Fry disponível nesse link. Estudos disciplinares consistem em pesquisas que se concentram em um objeto específico dentro dos limites de uma única disciplina acadêmica. Estudos multidisciplinares envolvem diferentes disciplinas acadêmicas investigando um tema ou problema sob um guarda-chuva temático. O resultado do conhecimento também é disciplinar e a cooperação entre as disciplinas é limitada. Já os estudos interdisciplinares envolvem duas ou mais disciplinas não relacionadas entre si que são postas em uso para investigar um mesmo objeto, de modo que pesquisadores cruzam as fronteiras entre disciplinas a fim de criar novos conhecimentos e teorias que vão além do escopo de cada disciplina isoladamente. E, por fim, os estudos transdisciplinares reúnem não apenas disciplinas acadêmicas, mas também participantes não acadêmicos e suas visões de mundo e conhecimentos, em direção a um objetivo de pesquisa e ação comum. Acho importante mencionar que, do meu ponto de vista, não há nenhum juízo de valor sobre essas abordagens. Penso que todas são relevantes e têm seu lugar e contribuição. Há beleza tanto no estudo de caracteres específicos de uma planta e sua relação evolutiva com outras espécies, como também na colaboração entre ciência e conhecimento botânico de comunidades indígenas e locais.
Voltando ao meu caso, trabalho com conservação de polinizadores e, mais recentemente, comecei a trabalhar com áreas urbanas. Considerando o primeiro caso, esse tema pode ser abordado de várias maneiras, por exemplo, testando como as práticas agrícolas influenciam a abundância e a diversidade de polinizadores. Temos evidências de que os polinizadores estão em declínio devido à agricultura, às mudanças climáticas, à urbanização, ao uso de pesticidas etc. Também sabemos que há uma falta significativa de dados a esse respeito em muitas partes do mundo. Parte da solução para abordar esse problema requer o envolvimento das pessoas na conservação de polinizadores, seja por meio de ações diretas de conservação seja por conscientização e elaboração de políticas públicas que melhor protejam esse serviço, por exemplo. No entanto, como podemos envolver efetivamente as pessoas na conservação de polinizadores?
Esta é uma situação típica em que pesquisadores/as da Ecologia precisam cruzar as fronteiras de sua disciplina para encontrar outros pesquisadores em outros campos igualmente disciplinares. Em minha pesquisa de doutorado, recorri teoricamente à psicologia social e metodologicamente às ciências sociais para entender aspectos sociais relacionados à conservação de polinizadores. Vamos explorar alguns dos desafios que encontrei ao conduzir pesquisas inter- e transdisciplinares, os quais merecem grande atenção de quem busca realizar estudos dessa natureza.
Um desafio significativo para o emprego de abordagens inter- e transdisciplinares em ciências ambientais é a dependência epistêmica. Este conceito está relacionado ao fato de que, em atividades de pesquisa colaborativa – como as interdisciplinares -, os cientistas que trabalham em um campo distante de sua formação acadêmica são dependentes epistemicamente de outros cientistas vinculados a este campo para compartilhar ideias, aprender e aplicar métodos e interpretar resultados. Em segundo lugar, na pesquisa inter- e transdisciplinar, geralmente o delineamento experimental ou amostral e a coleta de dados funcionam de maneira bastante diferente, notadamente quando integramos ciências sociais e ciências ambientais. Geralmente, métodos qualitativos se encontram com métodos quantitativos para dar sentido aos resultados e o estilo de escrita e comunicação são bastante diferentes. Além disso, trabalhar com esse tipo de dados requer o desenvolvimento de novas habilidades éticas e a consideração da disponibilidade de outras pessoas para contribuir com sua pesquisa, o que pode levar muito tempo. Por último, mas não menos importante, atualmente os cientistas são avaliados principalmente por seu histórico de publicações. Os resultados de pesquisas inter- e transdisciplinares nem sempre são aceitos em revistas disciplinares, embora haja alguns exemplos de revistas de alta qualidade que se concentram nesse tipo de pesquisa. Mas o mais importante é que manter a excelência na pesquisa – propondo questões que avancem o conhecimento, executando os estudos com rigor metodológico etc. – implica uma aprendizagem profunda de um campo completamente novo, o que traz desafios epistêmicos e linguísticos.
Do ponto de vista pessoal, eu adicionaria que lidar com algo desconhecido tanto para a minha formação acadêmica, quanto para a comunidade científica ao meu redor, representou um desafio ainda maior e, muitas vezes, me levou a ser questionada – principalmente por outros professores e colegas – se estava no lugar certo. Não posso deixar de mencionar que um questionamento externo só nos atravessa e nos marca quando há também um questionamento interno; afinal, nunca damos muita atenção a questões que já não existem em nós. No meu caso, como mulher negra fazendo algo diferente, essas questões também surgiram internamente, pois à época não tinha referências e experiências muito concretas que me inspirassem ou me acolhessem nesse caminho. Assim, a necessidade (e vontade) de me manter firme e inovar foram muito importantes nesse caso. Superar esse desafio requer resiliência e disposição. A criação de redes de contatos com pessoas abertas a abordagens inter- e transdisciplinares também é igualmente relevante.
A pesquisa inter- e transdisciplinar também oferece inúmeras oportunidades para ecólogos e conservacionistas. Em primeiro lugar, trabalhar em ambientes colaborativos e diversos promove o desenvolvimento de habilidades eficazes de comunicação, permitindo que os pesquisadores adaptem sua linguagem a diferentes públicos. Em segundo lugar, proporciona oportunidades para criar soluções baseadas na natureza e nas pessoas. No caso da pesquisa transdisciplinar, diversos conhecimentos são combinados. Isso pode influenciar diretamente a tomada de decisões por meio da participação de múltiplos atores, levando em consideração também a ciência. Por fim, as abordagens inter- e transdisciplinares permitem explorar como métodos e projetos de pesquisa de diferentes disciplinas podem ser combinados para informar esforços de conservação. Apesar dos desafios, a prática contínua e o engajamento com abordagens inter- e transdisciplinares reduzem gradualmente a percepção de estar “fora do seu campo”.
Algumas discussões adicionais sobre esse tema ainda estão em aberto, principalmente se considerarmos que há esse novo perfil profissional em ascensão que ocupa um não-lugar na academia como a conhecemos hoje, mas que também transforma este não-lugar em algo inventivo. Assim, precisamos refletir sobre como os ecólogos disciplinares ensinarão a uma nova geração de pesquisadores inter- e transdisciplinares? Essa nova geração realmente pode trazer mudanças significativas? Como os ecólogos básicos e aplicados podem se engajar efetivamente nesse diálogo? Qual é o estado atual do conhecimento inter- e transdisciplinar em Ecologia e Conservação? Que habilidades outras ainda precisaremos desenvolver para atuar na pesquisa inter- e transdisciplinar? As discussões atuais também destacam a importância de considerar as relações geopolíticas, particularmente entre o Norte e o Sul Global, por meio das quais países da Europa e América do Norte têm sistematicamente oprimido e dominado países da América do Sul e África, por exemplo. Também não podemos esquecer da necessidade de descolonizar a Ecologia e a Conservação, ou seja, de repensar a predominância do pensamento europeu nas soluções aos problemas socioambientais e abandonar práticas que reforçam injustiças ambientais. Autonomia, curiosidade e coragem são características necessárias para superar os desafios associados à formação em pesquisas inter- e transdisciplinares. Essa abordagem oferece um caminho promissor para construir um pensamento inovador e crítico sobre problemas socioambientais, além de promover a capacitação de futuros líderes e cientistas comprometidos em encontrar soluções criativas em um mundo em rápida mudança.
Caren Queiroz Souza Pesquisadora de Pós-doutorado Universidade Federal de São Carlos Brasil
A relação entre as florestas e as chuvas é conhecida por diversas culturas. A ciência vem descrevendo essa relação cada vez com mais detalhes. Mas existem teorias que explicam essa relação?
A teoria da bomba biótica de umidade explica que, por meio da transpiração e da condensação, as florestas criam ativamente regiões de baixa pressão, que absorvem o ar úmido dos oceanos, gerando ventos capazes de transportar umidade e sustentar a chuva nos continentes. Como as massas de terra continental estão acima do nível do mar, por efeito da gravidade toda a água líquida acumulada no solo e nos reservatórios subterrâneos flui inevitavelmente para o oceano, na direção da inclinação máxima das superfícies. Então, para acumular e manter reservas ideais de umidade na terra, seria necessário compensar o escoamento gravitacional de água para o oceano, por meio de um fluxo reverso de umidade, do oceano para a terra. Segundo essa teoria, esse fluxo reverso é impulsionado e mantido por grandes áreas contínuas de floresta. Isso quer dizer que, se a floresta for removida, o continente terá muito menos evaporação do que o oceano contíguo – com a consequente redução na condensação –, o que determinará uma reversão nos fluxos de umidade, que passarão a ir da terra para o mar, criando um deserto onde antes havia floresta. Por sua vez, ações de restauração florestal podem aumentar a precipitação local e também contribuir para o fortalecimento do transporte total de umidade do oceano para a terra continental, aumentando a magnitude e a confiabilidade da precipitação.
A Floresta Amazônica mantém o ar úmido em seu interior e exporta rios aéreos de vapor, que contribuem para formação de chuvas fartas e irrigam regiões distantes no verão do hemisfério sul. Nos últimos anos, os rios voadores, como têm sido chamados esses cursos de água atmosféricos, apareceram em matérias da mídia de grande circulação, a exemplo da BBC News Brasil e Revista Galileu. Eles foram definidos em 1992 como grandes volumes de vapor d’água que são transportados na baixa atmosfera. Um dos rios voadores de maior importância para a América do Sul é formado pela ação conjunta da forte evaporação das áreas tropicais mais quentes do Oceano Atlântico e da Floresta Amazônica. Sob ação da bomba biótica de umidade, a intensa evaporação nas áreas do oceano é sugada para dentro do continente e avança no sentido oeste, até atingir a Cordilheira dos Andes. Ao longo dessa trajetória, o vapor d’água recircula, tendo seu volume aumentado ao passar por cima da Floresta Amazônica, graças à atividade de evapotranspiração das árvores. Ele segue então seu caminho e desagua em áreas mais remotas e mais áridas nas regiões Sudeste e Sul do Brasil, além de dispersar-se pelos países fronteiriços, como Paraguai e Argentina.
O processo de evapotranspiração é a perda de água pelas plantas na forma de vapor. Esse processo se inicia pela ação de diferentes fatores, em especial pela mudança de temperatura na atmosfera, que interfere no movimento da água que circula pelo corpo da planta e chega à superfície das folhas. Depois que as nuvens precipitam, grande parte da água atravessa o dossel e se infiltra pela floresta. Parte dessa água é armazenada no solo, ou mais abaixo, nos aquíferos. A água que é armazenada no solo retorna para a atmosfera quando é absorvida pelas raízes e posteriormente liberada através da evapotranspiração das árvores. Essa água absorvida pelas raízes ascende para as folhas pelo xilema, tecido condutor de água e sais minerais nas plantas vasculares.
Apesar de a transpiração ocorrer em qualquer parte do organismo vegetal acima do solo, a maior proporção ocorre nas folhas (mais de 90%), o que está associado à sua anatomia. Na superfície foliar, existe uma camada de cera interrompida por poros, denominados estômatos. No interior desses poros, há células agrupadas que emitem vapor d’água para o espaço presente entre elas, no qual há uma abertura para atmosfera, o que possibilita a perda desse vapor, fenômeno que se conhece como evapotranspiração. À medida que a água vai sendo perdida para a atmosfera, o seu movimento pelo corpo da planta continua garantindo a hidratação dos tecidos internos e a manutenção do próprio ciclo de evapotranspiração. Para que exista essa elevação da coluna d’água pelo tecido condutor, as moléculas de água precisam estar coesas e submetidas a uma força de tensão que vence a gravidade. Essa dinâmica no transporte da água do solo para as raízes e das raízes para o caule depende da pressão positiva de raiz e da capilaridade. A pressão positiva de raiz resulta da constante perda de água do interior do corpo da planta, criando uma força de arrasto da água do solo em direção às raízes, que acaba elevando a coluna d’água pela extensão do xilema, mas com um poder de ascensão limitado em função da força da gravidade, que se opõe a essa elevação. A capilaridade contribui, então, para esta ascensão da coluna d’água. Trata-se de um fenômeno físico que ocorre quando líquidos se deslocam na superfície de tubos muito finos, o que depende das propriedades de coesão e adesão. A força de adesão depende da afinidade existente entre o líquido e a superfície sólida do tubo. Já a força de coesão depende da atração entre as moléculas do próprio líquido e atua no sentido oposto à parede do tubo. Como a água está sendo perdida pela evapotranspiração, as folhas atuam como uma bomba de sucção que mantém este processo funcionando.
No relatório O futuro climático da Amazônia, Nobre revelou que, usando dados de evaporação coletados nas torres de fluxo de um projeto de grande escala, foi possível estimar a quantidade total diária de água fluindo do solo para a atmosfera através das árvores na bacia amazônica. O valor estimado, para uma área de 5,5 milhões de km2, chegou ao total surpreendente de 20 bilhões de toneladas de água transpiradas ao dia para a atmosfera (ou 20 trilhões de litros). Se todas as florestas da porção equatorial da América do Sul fossem consideradas, esse número passaria a 22 bilhões e, se considerássemos as florestas que existiam em 1500, seriam 25 bilhões de toneladas ou mais. Para efeito comparativo, o rio Amazonas despeja no Oceano Atlântico cerca de 17 bilhões de toneladas ao dia, pelo menos 3 bilhões de toneladas a menos do que foi estimado na formação do rio aéreo.
Mais um fenômeno controlado pela floresta e capaz de atuar no clima diz respeito às emissões de aerossóis moduladas pelas árvores. Esses aerossóis são elementos-chave do sistema climático, pois são capazes de mudar os padrões de chuva na região amazônica, como consequência da redistribuição de energia e da formação de núcleos de condensação e núcleos de gelo. Os aerossóis controlam a formação de nuvens e a precipitação através de seus efeitos sobre os núcleos nos quais gotículas de água se condensam ou gelo se forma. Nuvens são agregados de gotículas em suspensão no ar. Em baixas temperaturas, estas gotículas se condensam a partir do vapor. Porém, para formar núcleos de condensação, é preciso haver uma superfície sólida ou líquida que funcione como “semente” para que se inicie a deposição e condensação das moléculas de vapor. Essas sementes são geradas pelos aerossóis encontrados na atmosfera. A depender de sua composição e abundância, eles podem espalhar ou absorver radiação, assim como aumentar ou suprimir a precipitação.
Os aerossóis são classificados em partículas primárias, produzidas deliberadamente pela flora (por exemplo, liberação de pólen e esporos de fungos) e incidentalmente (por exemplo, como restos de folhas e solo ou como microorganismos em suspensão), e partículas secundárias, produzidas na atmosfera pela oxidação de gases residuais, que resulta em compostos de baixa volatilidade. Assim como partículas de aerossóis de outras origens (por exemplo, poeira mineral, sal marinho, fumaça de biomassa – oriunda da queima de biomassa – e partículas de poluição), partículas biológicas podem influenciar a formação de nuvens e processos de precipitação através de diversos mecanismos, os quais são cruciais para a manutenção do ciclo hidrológico. A precipitação induzida por partículas primárias e secundárias, emitidas pelas florestas ou formadas na atmosfera, agindo como núcleos de condensação ou núcleos de gelo, sustenta a reprodução de plantas e microrganismos no ecossistema do qual os precursores dessas partículas são emitidos (Figura 1). Essa causalidade circular é retratada na pergunta feita por Nobre em seu texto de 2007: “As florestas crescem onde chove ou chove onde crescem florestas?” Ela pode ser entendida como um dilema do tipo ovo-ou-galinha e estimular a seguinte conclusão: “[…] onde tem mata, tem chuva” (Nobre, 2007, p. 369).
Há diferentes formas de interpretar a causalidade circular entre florestas e chuvas. Uma delas é uma proposta recente que vem sendo discutida na filosofia da biologia: a teoria organizacional das funções ecológicas. Na próxima seção, falaremos sobre ela e a abordagem que a fundamentou, a teoria da autonomia biológica.
Figura 1: Principais mecanismos associados ao controle da água pela floresta no ciclo hidrológico continental. A água presente no solo entra no corpo da planta, sendo em seguida conduzida até as folhas, onde grande parte é evapotranspirada. As árvores também contribuem com compostos orgânicos voláteis, que oxidam em contato com a atmosfera e são os maiores responsáveis pela formação de núcleos de condensação. As partículas primárias, como esporos de fungos e grãos de pólen, contribuem para a formação de núcleos de gelo, mas também podem agir como núcleos de condensação “gigantes”, gerando grandes gotas e induzindo chuva quente, sem formação de gelo. Autor da figura: Jeferson Coutinho.
Sistemas biológicos: um tipo específico de regime causal
Uma forma de interpretar a circularidade causal entre organismos e componentes abióticos num ecossistema vem sendo explorada na construção de uma teoria organizacional das funções ecológicas. De acordo com essa teoria, assim como em outras abordagens, como a teoria da construção de nicho ou a teoria Gaia, a vida influencia as condições físico-químicas do ambiente de uma maneira que acaba por contribuir para a sua própria auto-manutenção. A teoria também compartilha com outras versões de abordagens organizacionais a ideia de fundamentar as atribuições de funções a componentes dos ecossistemas com base na proposição de que os sistemas biológicos realizam um tipo específico de regime causal, no qual as ações de um conjunto de partes são condição para a persistência de toda a organização ao longo do tempo. Essa teoria é derivada da teoria da autonomia biológica, que propõe que sistemas vivos são irredutíveis a sistemas físico-químicos, exibindo propriedades qualitativamente distintas porque apresentam uma organização específica, que pode ser descrita como um fechamento, isto é, uma circularidade causal diferente daquela encontrada em sistemas físico-químicos, como explicaremos a seguir.
A teoria da autonomia biológica considera que sistemas vivos são organizacionalmente fechados e termodinamicamente abertos. Quando dizemos que um sistema vivo é organizacionalmente fechado, estamos nos referindo ao que denominamos acima “circularidade causal”. Em termos mais precisos, isso significa que a organização biológica em questão (por exemplo, de um organismo ou ecossistema) exibe “fechamento”, isto é, seus componentes e suas operações dependem umas das outras para sua própria produção e manutenção, e determinam coletivamente as condições para que o próprio sistema exista e siga existindo. Conforme a teoria da autonomia biológica, o fechamento característico dos sistemas vivos é um “fechamento de restrições”. Isso requer, claro, que expliquemos o que são restrições.
Restrições são causas locais e contingentes, exercidas por estruturas e processos específicos, que reduzem os graus de liberdade da dinâmica ou do processo sobre o qual atuam, mas permanecem conservadas na escala de tempo relevante para descrever sua ação causal em relação àquele processo ou dinâmica. Como as restrições reduzem os graus de liberdade dos processos internos ao sistema vivo, elas contribuem para sua coordenação, a qual gera, por sua vez, novas possibilidades de comportamento e adaptação ao meio para o sistema como um todo. Uma variedade de entidades pode desempenhar o papel de restrições em um organismo, por exemplo, macromoléculas (digamos, enzimas ou ribossomos) e configurações materiais específicas (como o sistema de vasos sanguíneos ou os circuitos neurais dentro do cérebro). Reduzir graus de liberdade de um processo significa dizer que, sob a ação da restrição, o processo tem um universo menor de possíveis trajetórias, em comparação com o que teria na ausência da restrição. É essa redução de graus de liberdade que faz com que os processos sejam, sob a influência das restrições, mais coordenados, de tal maneira que a manutenção da vida seja possível. Portanto, quando nos referimos a um fechamento de restrições, estamos considerando uma rede de dependências mútuas entre partes constitutivas de um sistema que atuam como restrições, oque contribui não apenas para a manutenção e existência das outras partes do sistema, como também do próprio sistema como um todo e, consequentemente, de si mesmas.
De modo diferente dos sistemas vivos, cadeias circulares de processos também podem ocorrer sob a ação de restrições externas. Esse comportamento é tipicamente observado em sistemas físicos ou químicos e é caracterizado por sequências ordenadas de ocorrências ou estados dinâmicos que estão ligados sistematicamente uns aos outros, tipicamente de maneira causal. O fechamento de processos, como é chamado, ocorre quando esses estados ou ocorrências formam um ciclo fechado: um processo A causa um processo B, que causa um processo C, que, por sua vez, causa A. É o que ocorre, por exemplo, no fluxo circular que a água percorre sob ação da radiação solar em uma garrafa de vidro fechada e preenchida até a metade (Figura 2).
Figura 2: Fechamento de processos no fluxo circular que a água percorre sob ação da radiação solar em uma garrafa de vidro fechada e preenchida até a metade: (1) a radiação solar atravessa as paredes da garrafa e aquece a água; (2) ao atingir uma determinada temperatura, a água começa a evaporar; (3) o vapor d’água, depois de subir, condensa no topo da garrafa e cai como água líquida; (4) que fica novamente sujeita à evaporação. Autora da figura: Clarissa Leite.
A ciclagem das moléculas de água dentro da garrafa é um fluxo termodinâmico, físico-químico, circular, limitado apenas por entidades externas, a exemplo do vidro. O vidro atuam, então, como uma restrição externa, que não é regenerada pelo fluxo termodinâmico cíclico da água. No caso de um fechamento de processos, as restrições são apenas externas, não dependendo da dinâmica sobre a qual atuam, como no exemplo acima. Em vez de apenas uma cadeia circular de processos influenciada por restrições externas, sistemas biológicos produzem restrições internas, que atuam sobre seus próprios processos e, assim, exibem dois regimes causais distintos, mas interdependentes: um regime termodinâmico aberto de processos e reações e um regime fechado de dependência entre componentes que agem como restrições. Isso é o que significa dizer que esses sistemas são termodinamicamente abertos e organizacionalmente fechados.
De acordo com a teoria da autonomia biológica, funções são sempre atribuídas a componentes que atuam como restrições internas a um sistema vivo. Usando um exemplo clássico nos debates sobre funções, podemos considerar o coração para explicar o que significa atribuir função a uma restrição de acordo com essa teoria. O coração é uma parte constitutiva de muitos animais que tem a função de bombear sangue. Interpretado nos termos da teoria da autonomia biológica, pode-se afirmar que, ao bombear sangue, o coração atua como uma restrição. Primeiro, porque o bombeamento de sangue pelo coração tem o poder causal de alterar, por exemplo, a distribuição de gases e nutrientes num corpo animal, no sentido específico de que diminui os graus de liberdade desse processo de distribuição (o que ocorre, claro, com a contribuição de várias outras restrições, a exemplo de todo o conjunto de artérias, arteríolas, capilares etc.). Segundo, considerando-se que leva cerca de um minuto para o sangue circular por todo o corpo de um ser humano (a título de exemplo), podemos perceber que, na escala temporal em que o coração, ao bombear sangue, restringe a distribuição de gases e nutrientes no corpo, ele se mantém conservado (assim como as artérias, arteríolas, capilares), no preciso sentido de que ele não sofre alterações nas propriedades relevantes para sua atuação como restrição que sejam devidas ao processo de circulação do sangue, naquela escala temporal.
A atribuição de uma função a uma restrição, conforme essa teoria, se apoia exatamente no papel causal que um determinado componente de um sistema exerce, como restrição, em processos vitais de um sistema vivo, dentro do fechamento de restrições que caracteriza sua organização. Isso implica que esta parte contribui para a manutenção da organização do corpo e é, ao mesmo tempo, mantida graças ao papel de outras restrições e desta organização mesma. Apliquemos novamente isso ao caso do coração. Ao bombear sangue e, assim, atuar como uma restrição sobre a distribuição de gases e nutrientes, o coração contribui para a manutenção e existência de todas as outras partes do corpo, bem como do próprio organismo, o que, por sua vez, contribui para a manutenção e existência do próprio coração. Retomando algo que explicamos acima, o fechamento de restrições destaca exatamente essa característica dos sistemas biológicos: que seus componentes constitutivos e operações dependem uns dos outros para sua manutenção e, além disso, contribuem coletivamente para determinar as condições sob as quais o próprio sistema pode existir.
Na teoria da autonomia biológica, as restrições que são produzidas sob influência de outras restrições são chamadas de dependentes, enquanto aquelas que participam do processo de produção de outras restrições, são chamadas de possibilitadoras. Para fazer parte do fechamento de restrições, uma restrição deve ser tanto dependente, quanto possibilitadora. Mas, e nos casos em que as restrições são apenas dependentes ou apenas possibilitadoras? As restrições que são exclusivamente possibilitadoras ou dependentes estabelecem conexões entre o sistema vivo e outros sistemas (vivos ou não), que ou constituem seu ambiente, ou se situam em níveis físico-químicos de processos internos aos seres vivos. Isso quer dizer que, para caracterizar um sistema como organizacionalmente fechado, não há a necessidade de afirmar que todas as restrições que agem na dinâmica do sistema são parte do fechamento. Mostra também que falar de fechamento organizacional não implica defender alguma independência do sistema vivo em relação ao ambiente. Um sistema que realiza fechamento organizacional de restrições é um sistema fisicamente aberto, inerentemente acoplado ao ambiente com o qual troca energia e matéria. Sem essa conexão com outros sistemas e sem troca de matéria e energia, o sistema vivo não tem como se auto-manter.
A explicação organizacional da teoria da autonomia biológica vem sendo construída desde os anos 1990, focando sobretudo sobre células e organismos. Enzimas e órgãos, como o coração, foram alguns dos exemplos usados para explicar como partes de sistemas vivos assumem funções quando atuam como restrições (p. ex., enzimas catalisando reações em células e, assim, diminuindo seus graus de liberdade; e órgãos, como o coração, diminuindo os graus de liberdade de processos como a distribuição de gases e nutrientes em determinados organismos). Em 2014, três pesquisadores aplicaram a explicação organizacional proposta por essa teoria a sistemas ecológicos, argumentando que as funções que componentes de sistemas ecológicos desempenham em processos ao nível do ecossistema como um todo podem ser entendidas como efeitos precisos (diferenciados) de componentes bióticos (vivos) ou abióticos (inanimados) que atuam como restrições sobre fluxos de matéria e energia nos ecossistemas. Isso demanda que os ecossistemas exibam uma organização que seja entendida em termos de um fechamento de restrições. Nunes-Neto, Moreno e El-Hani usaram para desenvolver a teoria o fitotelma de uma bromélia como modelo de um sistema ecológico organizacionalmente fechado, considerando as relações de predação e decomposição estabelecidas em uma teia alimentar interna a ele, envolvendo uma espécie de aranha, larvas de mosquitos e microorganismos, o que resulta numa diminuição dos graus de liberdade do fluxo de átomos como os de nitrogênio. O modelo foi representado com dois níveis hierárquicos, um relativo ao fluxo dos átomos e outro relativo aos papéis dos componentes bióticos que atuam como restrições. Essa proposta original deu os primeiros passos para a construção de uma teoria organizacional das funções ecológicas, que vem sendo aprimorada desde então.
Teoria organizacional das funções ecológicas e a formação de nuvens nos oceanos
Alguns anos depois, El-Hani e Nunes-Neto abordaram a transição de um mundo pre-biótico – composto de sistemas puramente físico-químicos – para um mundo controlado pela vida, com base na teoria organizacional. Eles descreveram como o sistema de formação de nuvens nos oceanos não resulta apenas de uma sequência de eventos físico-químicos relacionados à evaporação e precipitação da água (i.e., não resulta apenas de um fechamento de processos). Há uma participação ativa e decisiva de uma rede de interações de organismos marinhos, em especial do fitoplâncton, que levam à secreção de uma substância sulfurosa, o dimetilsulfureto (DMS), que contribui para a formação de núcleos de condensação de nuvens sobre o oceano, em um processo semelhante àquele que ocorre no continente envolvendo aerossóis liberados pelas florestas, tal como explicamos acima. Quando as nuvens precipitam, a chuva traz para o oceano precursores do dimetilsulfureto, que se tornam disponíveis para o metabolismo dos organismos marinhos, fechando, então, o ciclo estabelecido entre eles e a formação de nuvens. Como observado por estes autores, é importante notar que existe uma relação de dependência mútua entre a microbiota marinha e as nuvens, que pode ser entendida em termos de sua atuação como restrições, em escalas temporais específicas, sobre processos físico-químicos e, mais especificamente, como restrições dependentes e possibilitadoras no controle que o sistema exerce sobre o fluxo do dimetilsulfureto. A microbiota depende do enxofre depositado pela precipitação das nuvens e carreado pelos rios, assim como as nuvens dependem do enxofre derivado do dimetilsulfureto produzido pelo fitoplâncton. Assim, os autores propõem que a teoria organizacional das funções ecológicas oferece uma fundamentação consistente para explicar a transição de um fechamento de processos, no qual os ciclos do enxofre e da água correspondiam somente a uma sequência fechada de estados dinâmicos físico-químicos, para um sistema caracterizado por um fechamento de restrições, no qual a vida passa a exercer controle sobre estes dois ciclos. Se generalizarmos esse argumento, podemos chegar, então, a uma tese central da teoria Gaia, a de que, quando os seres vivos passaram a controlar parte importante dos processos físico-químicos planetários, passamos de um mundo controlado somente por processos físico-químicos a um mundo controlado pela vida.
Nesse texto sobre a relação da vida marinha com a formação de nuvens nos oceanos, El-Hani e Nunes-Neto tratam da atribuição de funções ecológicas a componentes inanimados ou abióticos. No entanto, para incluí-los como itens funcionais, eles devem, assim como os itens bióticos, atender a um critério fundamental da teoria: estar sujeitos ao fechamento, ou seja, atuar como restrições internas à organização do sistema e, portanto, sob seu controle. Como vimos anteriormente, ser uma restrição interna à organização do sistema quer dizer, segundo essa teoria, que o componente (nesse caso, abiótico) deve ser uma restrição dependente e possibilitadora. Caso não sejam restrições internas ao fechamento organizacional, não se pode atribuir funções ecológicas a esses componentes. Note-se, contudo, que eles ainda podem ser considerados relevantes na dinâmica do sistema ecológico, já que podem agir como restrições externas que afetam seus processos, mesmo não sendo parte de sua organização interna.
Explorando um outro caso em um texto ainda não publicado, os proponentes dessa abordagem explicam que, em ecossistemas savânicos, o controle exercido por espécies de plantas adaptadas ao fogo, por meio de características relacionadas à sua inflamabilidade, i.e., à facilidade com que entram em combustão, é um exemplo de como uma restrição anteriormente externa ao sistema (o próprio fogo) pode passar a fazer parte de sua dinâmica interna, quando colocada sob controle de sua organização. Quando o fogo é integrado à dinâmica do ecossistema por meio de espécies de plantas adaptadas a ele, que exibem características de inflamabilidade e influenciam sua frequência, ele passa a ter um papel construtivo na dinâmica do ecossistema. O fogo passa a ser uma restrição possibilitadora nos ecossistemas savânicos por seu papel nos processos de rebrotamento das plantas e é, ao mesmo tempo, uma restrição dependente, na medida em que sua produção depende, em parte, das espécies de plantas adaptadas a ele. Nesses casos, pode-se até falar em coevolução do fogo e da biota. Quando o fogo não está sob controle do sistema, ele não age como uma restrição funcional interna ao mesmo, mas apenas como uma restrição externa, que age sobre componentes do ecossistema, mas sem estar sob controle de sua organização, o que pode ser uma razão para que o fogo tenha um papel destrutivo.
Mas além da relação entre os organismos marinhos, a água e o enxofre nos oceanos, bem como das plantas com o fogo em ambientes savânicos, como a teoria organizacional das funções ecológicas poderia explicar a relação entre as florestas e a chuva nos continentes?
Teoria organizacional das funções ecológicas e o ciclo hidrológico continental
A partir dos argumentos apresentados acima, podemos refletir sobre o caso do ciclo hidrológico continental, o qual abordamos no começo desse texto. Seria este também um caso de fechamento organizacional de um sistema ecológico, no qual as árvores assumem papel funcional crucial nas trocas entre o continente e a atmosfera? Assim como no exemplo do coração, tomando como base a explicação organizacional, podemos entender que as árvores são parte constitutiva do sistema hidrológico nos continentes e têm a função de controlar trocas entre o continente e a atmosfera. Mais especificamente, elas restringem o fluxo de água, energia, carbono e outros elementos entre esses dois ambientes. Ao fazerem isso, elas contribuem para a manutenção e existência de outras partes (por exemplo, as nuvens, a água) do sistema hidrológico nos continentes, bem como para a manutenção e existência do próprio sistema e de si mesmas.
Além disso, podemos dizer que a água em florestas úmidas seria inicialmente uma restrição externa, mas teria sido recrutada, como restrição possibilitadora e dependente, para compor a organização interna desses ecossistemas, assim como no caso do fogo em ecossistemas savânicos? A água é considerada um dos principais agentes modeladores em florestas úmidas, afetando a distribuição, fisionomia e diversidade de espécies características desses ecossistemas. A água pode desempenhar, então, funções relevantes nos processos ecossistêmicos de uma floresta úmida, enquanto está sob o controle de restrições internas à sua organização, como as plantas. Por exemplo, características das plantas podem determinar diferentes volumes de evapotranspiração e modular a emissão de aerossóis nucleadores de nuvens nesses ecossistemas. Esses aspectos interagem com outros fatores do clima, determinam o volume de água que circula no sistema e influenciam o regime de chuvas no continente. Uma vez que a água precipita, sua participação na germinação das sementes é de fundamental importância. E são as plantas que germinam dessas sementes que então irão evapotranspirar e emitir aerossóis para a formação de mais chuva, fechando o ciclo.
Se a Floresta Amazônica e a água são restrições no fechamento organizacional do sistema hidrológico continental, podemos interpretar que elas determinam não apenas a manutenção e existência uma da outra, como também do próprio sistema. Sendo assim, a afirmação que podemos fazer, seguindo a teoria organizacional das funções ecológicas, é que as florestas crescem onde chove e chove onde crescem florestas. Assim como nos oceanos, em algum momento da história evolutiva da Terra ocorreu, nos continentes, a transição de um fechamento de processos para um sistema caracterizado por um fechamento de restrições, ou seja, para um mundo controlado pela vida em que as plantas passaram a atuar como agentes facilitadores, mas também dependentes do sistema hidrológico continental.
Clarissa Machado Pinto Leite (Instituto Nacional de Ciência e Tecnologia em Estudos Interdisciplinares e Transdisciplinares em Ecologia e Evolução – INCT IN-TREE, Universidade Federal da Bahia)
Jeferson Gabriel da Encarnação Coutinho (Instituto Federal de Educação, Ciência e Tecnologia da Bahia/IFBA, Instituto Nacional de Ciência e Tecnologia em Estudos Interdisciplinares e Transdisciplinares em Ecologia e Evolução – INCT IN-TREE, Universidade Federal da Bahia)
PARA SABER MAIS
Gorshkov, VG, Makarieva AM. Biotic Regulation: Main Page. Disponível em: <https://www.bioticregulation.ru/>. Acesso em: 21 de agosto de 2023.
<Nasa Global Tour of Precipitation in Ultra HD (4K) – Youtube. 2016. Youtube NASA Goddard, 20 de maio de 2016. Disponível em: <https://www.youtube.com/watch?v=c2-iquZziPU/>. Acesso em: 21 de agosto de 2023.
Nobre AD, 2014. O Futuro Climático da Amazônia, Relatório de Avaliação Científica. Patrocinado por ARA, CCST-INPE, e INPA. São José dos Campos, Brasil, 42p.
Pearce, F. 2020. Weather Makers. Science (American Association for the Advancement of Science) 368.6497: 1302-305. Acesso em: 21 de agosto de 2023.
A modelagem de sistemas ecológicos requer passos que devem ser bem compreendidos por quem realiza a modelagem e por quem deseja entendê-la. A interface da Ecologia com a Filosofia contribui para o esclarecimento de conceitos, além de cumprir um papel heurístico na construção de processos de modelagem mais profundos e inteligíveis.
Da interação interdisciplinar, emergem entendimentos e novos questionamentos. Autor: Jeferson Coutinho.
Como o encontro entre uma filósofa da ciência e um ecólogo pode contribuir para o avanço de uma área da Ecologia? Temos algo a ganhar com esse diálogo? E se a resposta for “sim”, quem exatamente ganha nessa história? Foi na busca de uma solução para os problemas existentes em determinada área da Ecologia que um tal diálogo se iniciou, envolvendo o ecólogo Jeferson Coutinho e a filósofa da ciência Luana Poliseli. A questão central que impulsionou esse diálogo foi a tentativa de modelar conceitualmente sistemas ecológicos para entender mais claramente quais seriam as variáveis ambientais que mais influenciariam a diversidade funcional das abelhas nos agroecossistemas e suas repercussões para o serviço de polinização. Mais precisamente, a intenção era saber como a diversidade de atributos morfológicos, fisiológicos e comportamentais das abelhas poderiam afetar o serviço de polinização em sistemas agrícolas e como um sistema agrícola poderia funcionar como um filtro capaz de selecionar o conjunto de tais atributos disponíveis nesse tipo de sistema.
Esta questão central levantou várias outras questões acessórias que necessitavam de aprofundamento teórico para uma melhor compreensão do fenômeno de interesse. Algumas dessas questões foram: Como medimos a diversidade funcional em Ecologia? Que conceito de função usamos em estudos empíricos? Como entendemos a relação entre variáveis ecológicas em diferentes escalas espaço-temporais? Todas essas questões exigiram uma exploração minuciosa do que já era conhecido na literatura ecológica, mas também demandaram análises da consistência teórica do que estava disponível para se trabalhar.
Notavelmente, também precisávamos encontrar uma maneira de modelar o sistema de interesse por meio de etapas pragmáticas, à luz de uma teoria da explicação consistente com o que estávamos procurando. Diante das necessidades expostas, a interação com Luana, com sua formação e experiência em filosofia da ciência, abriu campos de diálogo muito frutíferos, em particular se tivermos em vista a grande complexidade do sistema em análise. O primeiro avanço foi uma compreensão mais clara do que estávamos tentando analisar. No processo de modelagem teórica, é fundamental termos clareza sobre o que queremos modelar, apontando o contexto relevante do fenômeno. Nos encontros realizados, os questionamentos feitos por Luana serviram como balizadores para minha própria compreensão do fenômeno, pois era inevitável o uso de exemplos didáticos e analogias para uma melhor compreensão do que eu queria dentro da Ecologia. A cada nova indagação, o uso de recursos imagéticos e esquemáticos (Figuras 1 e 2) foi fundamental para estabelecer maior clareza sobre o que buscávamos modelar, requerendo explicações dos mecanismos subjacentes ao fenômeno de interesse, buscando-se compreender cada vez mais quais níveis de análise, ou seja, quantos e quais níveis mecanísticos e contextuais seriam importantes em nosso caso particular.
Figura 1: Primeiro esboço dos mecanismos subjacentes ao fenômeno que estávamos interessados em modelar. Nesse primeiro momento, havia noção de 3 escalas espaciais de interesse na modelagem do fenômeno ecológico, que tinha a intenção de propor um modelo preditivo sobre a estrutura funcional de uma comunidade de abelhas nativas, bem como acerca da manutenção de seus serviços de polinização em um agroecossistema. Do lado esquerdo, indicamos que há dinâmicas no nível das metacomunidades, na mancha de hábitat e no nível das flores individuais que são importantes filtros ambientais que determinam a estrutura funcional das comunidades de abelhas. Nesse momento, ainda não estavam claras as possíveis relações dentro de cada escala de observação (lado direito do diagrama), o que apontou para um caminho de busca de entendimento de relações complexas entre níveis de organização, escalas de observação, bem como fatores reguladores de populações e comunidades, para que houvesse melhor entendimento do fenômeno de interesse.
Figura 2. Esboço dos mecanismos que ocorrem na escala da paisagem, considerando três dos seus descritores: proporção de agricultura, diversidade e configuração. Para cada descritor, são propostas previsões da diversidade funcional de abelhas em agroecossistemas, de acordo com os modelos de metacomunidades. A emergência do entendimento sobre esses mecanismos e a proposição dessas previsões se deram com o uso das heurísticas descritas no corpo do texto e ao longo de constantes interações com Luana Poliseli. Mais detalhes sobre os modelos de metacomunidades podem ser vistos em Leibold et al. (2004). Algumas dessas previsões sobre a configuração da diversidade funcional em agroecossistemas foram testadas e os resultados apresentados no artigo de Coutinho et al. (2021).
Nesse processo, Luana trouxe constantes contribuições à medida que se debruçava sobre meu próprio progresso na compreensão do fenômeno e a minha forma de estabelecer os pilares da modelagem. Detalhes sobre esta análise podem ser vistos em artigo publicado no ano de 2020, no qual ela apresenta um modelo sobre a emergência da compreensão científica, que expõe as principais características dessa compreensão, como sua formação gradual, sua relação entre habilidades e imaginação, e sua capacidade de seletividade do conhecimento disponível.
Contudo, entender o que queríamos modelar foi apenas o primeiro passo. Ao longo de nossas interações, sentimos a necessidade de utilizar uma teoria da explicação científica, apoiando-nos em entendimentos relativamente recentes sobre o uso de explicações mecanísticas na ciência, como podemos ver, por exemplo, em trabalho de John Matthewson e Brett Calcott. Essa escolha decorreu das possibilidades que essa abordagem nos deu para modelar fenômenos que envolviam processos operando em múltiplas escalas de espaço e tempo, bem como da perspectiva de que relações não-lineares aconteciam entre processos ocorridos nessas escalas. Pautamos a escolha em um estudo aprofundado e colaborativo da literatura da chamada nova filosofia da ciência mecanística, guiada por discussões que visavam entender a abordagem e como o fenômeno ecológico poderia ou não ser modelado de acordo com ela. Nesse processo dialógico, percebemos que o fenômeno a ser modelado era de grande complexidade e, consequentemente, uma explicação mecanística não poderia ser totalmente aplicada ao nosso fenômeno, ainda que pudesse – como o fez – desempenhar um papel heurístico, ou seja, de orientação na condução da modelagem dos processos relevantes para o entendimento do fenômeno de interesse.
O novo corpo de conhecimento produzido foi um conjunto de heurísticas construídas de acordo com a nova filosofia da ciência mecanística e com teorias ecológicas. Essas heurísticas tiveram papel fundamental na pesquisa, pois guiaram a atividade de modelagem teórica em Ecologia. Este conjunto de heurísticas é descrito e discutido no artigo Philosophy of science in practice in ecological model building(Filosofia da ciência na prática na construção de modelos ecológicos). Nesse trabalho, discutimos o desenvolvimento de uma estrutura conceitual que unifica a explicação mecanística, as ciências dos sistemas complexos e a teoria de metacomunidades para modelar a estrutura funcional de uma comunidade de abelhas nativas, bem como a manutenção de seus serviços de polinização em um agroecossistema. Além disso, empregamos a teoria organizacional das funções ecológicas para esclarecer os conceitos e processos mais elementares a serem entendidos em termos funcionais ao longo de nosso trabalho, bem como a busca de variáveis, escalas espaço-temporais e relações de causa e efeito que pudessem apoiar a modelagem do fenômeno ecológico.
As seguintes heurísticas tiveram papel central nesse trabalho interdisciplinar de modelagem ecológica:
1) Caracterização do fenômeno: a descrição do fenômeno a ser modelado e do que será considerado em sua explicação;
2) Esboço do mecanismo: o desenvolvimento de diagramas (geralmente incompletos e passíveis de mudança ou mesmo abandono ao longo do processo de modelagem) para representar as relações entre os referenciais teóricos e o fenômeno, como podem ser vistos nas Figuras 1 e 2;
3) Estrutura hierárquica: esta heurística permite visualizar a interação entre diferentes escalas espaciais e temporais, criando uma estrutura que identifica e localiza os (e a quantidade de) níveis em que o mecanismo (ou mecanismos) estão organizados na superestrutura do fenômeno;
4) Condições possibilitadoras: as variáveis envolvidas nas atividades dos mecanismos sob investigação que são relevantes para a produção do fenômeno de interesse;
5) Distinção de componentes operacionais: essa heurística distingue os componentes e as funções das condições possibilitadoras dentro dos mecanismos envolvidos e especifica as relações e os limites entre esses componentes;
6) Mudança nos componentes operacionais: esta heurística permite explorar cenários alternativos e prever possíveis cursos distintos do sistema sob investigação, uma vez que os componentes operacionais sejam modificados;
7) Frequência de evidências: indica a causalidade entre os elementos das condições possibilitadoras, conforme informações probabilísticas e mecanísticas já disponíveis na literatura científica ou obtidas por meio das heurísticas anteriores, apontando, também, o que ainda carece de validação empírica;
8) Esquema do mecanismo: modelo mecanístico final, obtido após a utilização das heurísticas acima.
Esse conjunto de heurísticas auxiliou em diferentes momentos da elaboração do modelo teórico. Na primeira proposição do “esboço do mecanismo”, por exemplo, percebi que não conseguiria chegar a um esquema minimamente razoável de comunicação sobre o fenômeno e as variáveis relevantes. A cada novo esboço de mecanismo, novos desafios e novas soluções foram se apresentando, o que exigia mais esforço para compreender o que estava posto na literatura científica sobre o tema, indicando suas limitações e agregando as evidências que já estavam disponíveis, bem como buscando esforços de sínteses das teorias e dos conceitos que foram mobilizados no trabalho. A heurística “estrutura hierárquica” também contribuiu para a delimitação de processos que interagem em diferentes escalas de espaço e tempo e permitiu uma compreensão adequada do fenômeno sob modelagem.
A cada novo passo na construção do modelo, eu precisava revisar a literatura ecológica de uma perspectiva mais crítica sobre o fenômeno. Não se pode construir um modelo conceitual se não se conhece o conjunto de variáveis e as condições de interação entre elas que são relevantes para sua proposição. Isso não significa que devamos começar a proposição do modelo conhecendo todas as variáveis relevantes. Isso tiraria uma das grandes virtudes da arte de modelar: um refinamento gradual de nossas construções teóricas sobre o funcionamento do mundo. À medida que avançava na construção do modelo, mais inteligível o fenômeno se tornava. Foi possível estabelecer com mais clareza relações de causa e efeito de forma não espúria, na medida em que as heurísticas me colocavam em uma posição de investigação cada vez mais profunda acerca do fenômeno.
Os modelos derivados da teoria de metacomunidades, por exemplo, apontavam para processos ecológicos que interagem em diferentes escalas espaço-temporais, o que me ajudou a pensar com mais clareza sobre o sistema de interesse, elencando o que parecia mais fundamental para prever os rumos da diversidade funcional de abelhas no cenário modelado. Esta etapa foi fundamental para derivar novas hipóteses que poderiam ser testadas em estudos empíricos futuros.
Neste trabalho, estabelecemos a construção de uma narrativa unificacionista envolvendo um conjunto relevante de processos ecológicos e os potenciais padrões de diversidade que eles podem gerar no contexto dos agroecossistemas. Vejo isso como um avanço no campo da Ecologia, pois conseguimos olhar para o sistema de interesse de forma mais holística, o que também auxilia em propostas de gestão voltadas para a conservação dos processos ecológicos e dos padrões de diversidade que eles geram no espaço.
Dentre os importantes aprendizados derivados dessa pesquisa colaborativa, posso destacar: 1) a interação com a Filosofia da Ciência contribuiu para uma compreensão cada vez mais clara do fenômeno ecológico, na medida em que os questionamentos gerados me colocaram em uma posição de busca profunda por questões não tão claras na Ecologia Funcional; 2) a Teoria organizacional das funções ecológicas indicou caminhos mais robustos de delimitação conceitual na construção do arcabouço teórico, especialmente no que diz respeito à identificação de características ou traços das abelhas que têm importância funcional, os quais podiam ser traços de resposta ou traços de efeito (para mais detalhes a respeito desses dois tipos de traços, veja aqui); 3) o próprio arcabouço teórico contribuiu para a derivação de novas hipóteses que podem ser testadas em estudos empíricos; e 4) aprender a construir o modelo enquanto estava imerso nas etapas metodológicas de sua própria construção ajudou a refazer caminhos de compreensão do fenômeno ecológico, ressignificando a forma como concebo meu fenômeno de interesse.
Encorajo a utilização do conjunto de heurísticas que construímos nesse trabalho interdisciplinar, que aproximou Ecologia e Filosofia da Ciência, em outros estudos ecológicos, de modo que possamos investigar a adequabilidade dessas heurísticas em relação a outros fenômenos de grande interesse para a Ecologia. É importante ter claro, contudo, que não se trata de alguma espécie de receita, a ser meramente seguida, mas de um caminho derivado de uma interação frutífera que vem rendendo a pavimentação de uma linha de pesquisa pautada na interdisciplinaridade como forma de lidar com fenômenos complexos, que requerem múltiplos olhares na tentativa de torná-los mais inteligíveis.
Jeferson Gabriel da Encarnação Coutinho
Instituto Federal de Educação, Ciência e Tecnologia da Bahia (IFBA) e Universidade Federal da Bahia (UFBA)
As doenças são fenômenos comuns a todos os seres vivos. Quando elas se apresentam, o organismo mobiliza mecanismos internos que podem permitir manter uma adequada interação com o meio. Nesse sentido, ser saudável não significa só possuir um corpo saudável, mas também sentir-se bem quando agimos num meio socioambiental.
Um fato muito comum na vida dos seres vivos é que eles, às vezes, ficam doentes. Quando as doenças se apresentam, o organismo, por exemplo, o organismo humano, reage com mecanismos fisiológicos que, para remediar eficazmente a condição patológica, tentam ajustar o estado interno ao contexto ambiental externo. Isso permite que o individuo se sinta no melhor estado de saúde possível quando tiver de se relacionar novamente com este contexto.
Tomemos o exemplo de uma doença ocular chamada de “hemianopsia”. Nos casos em que esta doença se apresenta, a hemianopsia envolve a perda da metade do campo visual do sujeito. Uma completa hemianopsia direita, como aponta, por exemplo, David Hubel, implica o escurecimento do centro do campo visual: é como se, observando a palavra “céu” e tentando se concentrar no “é”, não se pudesse ver o “u”, mas apenas o “c” e o lado esquerdo do “e”.
Um aspecto interessante dos mecanismos regulatórios que podem ser observados quando esta doença surge foi salientado pelo filósofo da medicina francês Georges Canguilhem em “Visão como modelo do conhecimento”, um manuscrito inédito de 1956-57 que pode ser consultado, junto com outros textos inéditos de Canguilhem, na École Normale Supérieure de Paris.
Canguilhem salienta que, apesar da doença de que sofrem e, por conseguinte, do escurecimento total da parte central de seu campo visual, vários pacientes hemianópicos parecem não perceber nenhuma sensação de cegueira quando interagem com um interlocutor ou com os objetos externos.
Na verdade, um dos comportamentos típicos dos pacientes afetados por hemianopsia é o seguinte: ao invés de manter um olhar reto quando observam um objeto ou um interlocutor, eles olham de lado, mantendo a cabeça levemente reclinada. Nos sujeitos saudáveis, umas das partes do olho que permitem a visão se chama de fóvea. Como nos sujeitos hemianópicos a ação da fóvea está comprometida, o que se observa é que, em seus olhos, cria-se muitas vezes uma outra fóvea na metade intacta da retina, isto é, uma espécie de “nova fóvea”, que substitui a ação da fóvea danificada. Esta nova fóvea responde às necessidades do sujeito de focalizar os objetos externos ao seu redor. E o que é importante destacar é que a posição dela não é fixa, não é estática dentro da retina, mas muda de acordo com as necessidades e as intenções do sujeito, dependendo do ponto no espaço exterior que ele quer observar. Em suma, para restabelecer um certo estado de saúde, o organismo em questão lida com a patologia através de mecanismos fisiológicos que reorganizam os componentes orgânicos da retina, e isso a fim de manter, mesmo em condições anatomicamente anormais, uma interação eficaz com o contexto exterior.
São, portanto, o ambiente e a forma como um organismo específico interage com ele o que determina, entre outras coisas, os processos de adaptação e o estado de saúde dos organismos.
Estes argumentos de Canguilhem são similares a algumas teorias recentes da filosofia da biologia e da medicina, por exemplo, as teorias de Cristian Saborido e colaboradores sobre a mau-função orgânica. Saborido e colaboradores apontam que estabelecer o que é patológico num organismo, isto é, o que é uma “mau-função”, não deriva meramente de um cálculo estatístico, ou de uma simples observação dos estados fisiológicos do organismo. Estabelecer a existência de uma mau-função num paciente significa avaliar a relação concreta entre o organismo em questão e seu meio socioambiental. Por exemplo, uma síndrome de Gilbert não envolve nenhuma mau-função naqueles pacientes que se sentem saudáveis, na medida em que eles não percebem obstáculos durante as próprias atividades diárias (como sensações de cansaço), apesar de um aumento excessivo ou descontrolado da bilirrubina, um pigmento contido na bílis. Por um lado, a mau-função é algo objetivamente relacionado com a fisiologia de um organismo. Mas, por outro lado, se o organismo não perceber nenhum obstáculo durante sua interação com o contexto externo, pode-se dizer que ele não apresenta mau-funções e, por conseguinte, que ele é saudável.
Isso se deve àqueles mecanismos internos de regulação adaptativa aos quais Canguilhem também fazia referência. No caso da síndrome de Gilbert, estes mecanismos podem compensar com sucesso um excesso de bilirrubina, razão pela qual o sujeito se sente saudável durante sua interação com o meio. Em termos gerais, quando as condições ambientais mudam, o sistema adaptativo do organismo desencadeia ações para modular adequadamente o seu funcionamento. Neste caso, pode-se falar de regulação adaptativa, que é desencadeada pela forma como um ser vivo específico interage concretamente com um meio. Isso também é análogo ao que Canguilhem salientava relativamente ao processo generativo da “nova fóvea” que se desenvolve nos sujeitos hemianópicos. Para Canguilhem, com efeito, este tipo de fóvea surge (também) das necessidades ou intenções especificas de um indivíduo em um meio determinado, permitindo a este indivíduo agir adequadamente no meio e, assim, fazendo-o sentir-se saudável.
Ser saudável, como nos dizem os filósofos da medicina Canguilhem e Saborido, não significa apenas ter um corpo fisiologicamente saudável. Um corpo saudável previne tanto quanto possível a ocorrência de doenças, mas quando elas acontecem, o organismo desenvolve reações adaptativas internas que podem permitir manter uma interação adequada com o ambiente. Em resumo, ser saudável significa também sentir-se saudável.
Emiliano Sfara
(Pós-doutorando, Instituto de Biologia/UFBA)
PARA SABER MAIS:
Canguilhem, G. (1956-57) La vision comme modèle de la connaissance. Este manuscrito pode ser consultado no CAPHÉS (Centre d’Archives en Philosophie, Histoire et Édition des Sciences) em Paris, colocação: GC. 13. 2.
Canguilhem, G. (1966), O normal e o patológico, trad. port. 2009, Forense Universitária, Rio de Janeiro.
Mês de junho chegou ao fim e com ele acabou também o frenesi do “mês dos namorados”. Restaurantes lotados, propagandas em vermelho e rosa por todo lado e a pressão de escolher o presente perfeito. Essa não é nem de longe a descrição do cortejo mais esquisito do mundo animal.
Sexo é bom para os genomas e para as espécies. Como já discutido em um post anterior aqui no blog, a reprodução sexuada cria conjuntos inéditos de cromossomos por meio da combinação dos cromossomos parentais (recombinação) e permite reunir as mutações vantajosas e descartar as desvantajosas (permutação). Raramente, algumas espécies abrem mão do sexo e passam a apresentar algum tipo de reprodução assexuada. Nesses casos, as diferenças genéticas entre parentais e progênie ocorrem apenas por mutações. A maioria dessas linhagens estão destinadas à extinção. Um estudo recente testou essa predição ao examinar genomas de cinco espécies do bicho-pau do gênero Timema com reprodução exclusivamente por partenogênese e os comparou aos genomas de seus primos mais próximos com reprodução sexual. Os pesquisadores observaram que a partenogênese resulta em uma redução extrema da diversidade genética e muitas vezes leva a populações geneticamente uniformes. Também encontraram evidências de que a seleção positiva era menos efetiva em espécies partenogenéticas, sugerindo que o sexo é onipresente em populações naturais pois facilita taxas rápidas de adaptação. O estudo revela como a ausência de sexo afeta a evolução do genoma em populações naturais, fornecendo suporte empírico para as consequências negativas da partenogênese conforme previsto pela hipótese inicial. Este e outros exemplos mostram a importância do sexo para a manutenção da variabilidade genética, adaptação e sobrevivência das espécies. Já que existe sexo e ele é tão difundido, o sucesso evolutivo de uma estratégia de corte depende também do sucesso de atrair parceiros. Dada a importância evolutiva de ser atrativo, o investimento em um bom presente na hora do cortejo parece um pequeno preço a pagar perto dos benefícios de uma cópula bem-sucedida.
Em muitas espécies, as fêmeas somente copulam com machos que oferecem presentes nupciais. Esses presentes são de diferentes formas, dependendo da espécie. Na barata Xestoblatta hamata, as fêmeas se alimentam de secreções anais produzidas e “presenteadas” por seus parceiros após a cópula. Em outras espécies, as secreções não são tomadas oralmente, mas entregues juntamente com o esperma. Durante a cópula da mariposa Utetheisa ornatrix, o macho transfere um composto alcalóide para a fêmea para eventual incorporação nos ovos e a própria fêmea é a primeira beneficiária do alcalóide que recebe do macho. No final do acasalamento, o alcalóide recebido do macho já está geralmente distribuído por todo o corpo da fêmea (incluindo até as asas) e torna a fêmea protegida contra aranhas, que liberam a mariposa de suas teias devido a sua aversão ao composto. Um presente parecido é oferecido pelos machos da vespa Cosmosoma myrodora. Durante o cortejo, os machos recobrem as fêmeas de filamentos ricos em alcaloides com ação protetora contra aranhas. Em muitos casos, como na aranha Pisaura mirabilis, os presentes são presas nutritivas que a fêmea usa como recurso alimentar. Em casos mais extremos, o próprio macho é o presente nutritivo, como no caso do louva-deus e de muitas aranhas.
E quando aquele presente escolhido com tanto carinho não é bem o que o mozão estava querendo? Parece que isso está acontecendo com a baratinha, Blattella germanica. Machos dessa espécie oferecem às fêmeas um presente pré-acasalamento constituído de uma secreção rica em oligossacarídeos que inclui maltose e maltotriose, além de fosfolipídios, colesterol e vários aminoácidos. Ao atrair a fêmea para sua secreção altamente palatável, o macho coloca a fêmea na posição adequada para a cópula e, enquanto ela se alimenta da secreção nupcial, o macho ganha tempo suficiente para a cópula. Um estudo publicado em maio, mostrou, no entanto, que algumas fêmeas estavam recusando o presente nupcial (e as investidas do macho) e buscaram as causas desse novo comportamento.
Geralmente, as baratas adoram açúcar. Acontece que, devido ao uso de iscas tóxicas contendo glicose, as populações da baratinha desenvolveram rapidamente uma aversão comportamental adaptativa à glicose. Um artigo publicado em 2013 mostrou o mecanismo por trás dessa aversão em baratinhas (falamos rapidamente sobre esse exemplo em um post aqui no Darwinianas). As secreções das baratas macho têm diferentes tipos de açúcares –maltose e maltotriose, que geralmente são preferidas pelas fêmeas– assim como algumas gorduras. À medida que as fêmeas se alimentam de seu presente, a maltose é rapidamente convertida em glicose, e as fêmeas com aversão à glicose sentem um gosto amargo e param de se alimentar, o que também encerra a oportunidade de acasalamento. Os pesquisadores realizaram vários experimentos para verificar como a aversão à glicose afetava o cortejo das baratas. Eles descobriram que as fêmeas com aversão à glicose interrompiam a alimentação com maior frequência, especialmente quando o presente era oferecido por um macho selvagem (sem aversão à glicose). Os machos com aversão à glicose geralmente tinham níveis mais altos de maltotriose em suas secreções. A maltose é relativamente fácil de converter em glicose, enquanto a maltotriose é mais complexa e demora um pouco mais para se decompor em glicose e, portanto, machos com aversão à glicose tinham tempo extra para começar o acasalamento. Os pesquisadores também alteraram a qualidade da secreção masculina, substituindo as secreções de glicose e maltose por frutose. As fêmeas avessas à glicose desfrutaram de frutose e se alimentaram dela por mais tempo, resultando em sessões de acasalamento mais bem-sucedidas. No geral, a aversão à glicose evoluiu sob a seleção natural, favorecendo a sobrevivência na presença de iscas tóxicas, mas levando à interrupção da cópula. Assim, a seleção sexual está levando à modificação da secreção e comportamento sexual dos machos, que para garantir o sucesso da cópula, secretam mais maltotriose e copulam mais rapidamente.
Passamos do presente amargo para a propaganda enganosa. No caso da aranha P. mirabilis, o oferecimento de presentes nupciais pelo macho é mantido por seleção sexual. Os machos que presenteiam as fêmeas, têm uma taxa de 90% de sucesso na cópula, enquanto 40% dos machos sem os presentes conseguem copular. A duração da cópula é positivamente correlacionada com o tamanho do presente e a proporção de ovos fertilizados aumenta com o tempo de cópula. Assim, o presente nupcial é um bom investimento de recursos para o macho: induz a fêmea a copular, facilita o acoplamento e, ao prolongar a cópula, pode aumentar a quantidade de espermatozóides transferidos. O presente nupcial na espécie consiste em uma presa que o macho capturou e embrulhou em seda. O macho oferece esse presente e, se a fêmea aceitar o convite, ela agarra a presa embrulhada. Enquanto a fêmea está comendo, o macho realiza a transferência de esperma. Algumas vezes, no entanto, os presentes não são o que a fêmea esperava: os machos embrulham carcaças de animais que eles encontraram (ou comeram). Esses presentes não nutritivos são descobertos pela fêmea no meio da cópula, encurtando o período de transferência de esperma e aumentando a competição entre espermatozóides de machos diferentes. O oferecimento dos presentes “inúteis” tem um custo para esses machos e a escolha da tática de presente depende da disponibilidade de presas, de seu tamanho e tempo de maturação. Também existe o risco do próprio macho virar a refeição em vez de seu presente enganoso e, para fugir desse destino, alguns machos têm uma estratégia curiosa: eles se fingem de mortos ao apresentar seu presente para evitar o canibalismo. Assim que a fêmea começa a desembrulhar o presente, o macho volta à ação para copular.
Claro que os presentes dados no dia dos namorados não têm uma origem biológica comum aos presentes nupciais dos artrópodes. Nossos presentes (os chocolates, as flores) são resultado da nossa cultura e um prato cheio para o capitalismo, mas são oportunidades para reafirmar a atenção, investimento de tempo e recursos, algo que é realmente desejável em termos de cortejo humano. Quem não dispensa um chocolatinho, já tem um bom pretexto para garantir o do ano que vem: o ato de dar presentes é um comportamento ancestral cultivado pela seleção sexual! E quem não ficou satisfeito com o presente deste ano, pense que poderia ter sido bem pior.
Neste livro, a minha xará fictícia, a Dra. Tatiana, recebe cartas dos mais diversos animais e responde às dúvidas sobre corte, cópula e comportamentos pré- e pós-copulatórios. De uma forma bem divertida, explica a seleção sexual em seus conselhos aos animais aflitos.
Há muito tempo é de senso comum que o plástico é um grande problema para o meio ambiente. O microplástico vem ganhando cada vez mais atenção, principalmente no ambiente marinho. Embora pouco se saiba sobre o efeito do microplástico na saúde humana, recentemente um estudo mostrou que foram encontrados diferentes tipos de microplástico na corrente sanguínea. Se está interessado em saber um pouco mais sobre este assunto, seja bem-vindo, neste post falaremos a seu respeito.
Imagem de abertura: Fonte Pixabay (Pixabay License)
Já não é a primeira vez que idealizo escrever sobre um tema aqui no Darwinianas e mudo de ideia em cima da hora. Estava bolando um texto sobre os efeitos da Covid-19 na saúde mental, incluindo a perda de memória. Já tinha arrumado uma estrutura legal para o texto, pensado em relações com aspectos ecológicos, epidemiológicos e sociais… e eis que me deparo com uma matéria do The Guardian sobre a descoberta de microplásticos na corrente sanguínea de seres humanos. Mudança total de planos…
Vamos compreender um pouco mais sobre esse tema, que tem sido cada vez mais frequente em conversas e que está muito longe de ser divulgado no nível que merece. É importante avaliarmos o que podemos fazer, como podemos buscar soluções para um problema planetário tão amplo e desafiador. No entanto, para entendermos o tamanho do problema com o qual já estamos lidando, precisamos compreender um pouco melhor o que são os microplásticos e quais são suas origens. Assim, ficará mais claro o impacto desta descoberta.
A produção de plástico vem crescendo exponencialmente. Atualmente, são produzidas 400 milhões de toneladas anualmente, das quais apenas 12% são incineradas e 9% são recicladas, de acordo com relatório do Programa Ambiental das Nações Unidas (UNEP, sigla em inglês). O restante é descartado em aterros ou simplesmente lançado no ambiente, incluindo o oceano. Com isso, é esperado que o fluxo de plástico para os ambientes aquáticos triplique, passando de 11 milhões de toneladas em 2016 para 29 milhões de toneladas em 2040. É muita coisa! Todo plástico produzido sofre algum tipo de desgaste, seja durante o uso ou quando descartados no ambiente. É justamente a partir deste desgaste que fragmentos chamados de microplásticos, menores do que 5 mm de comprimento, são liberados e causam a poluição ambiental. As fontes de entrada dos microplásticos nos ambientes são muitas, incluindo cosméticos, roupas, embalagem diversas e processos industriais. O microplástico é classificado como primário, quando entra no ambiente já fragmentado, como é o caso de microfibras de roupas, por exemplo, ou como secundário, quando é fragmentado a partir de partículas maiores no ambiente, como exposto acima. Independentemente da origem e da classificação, sabe-se há muito tempo que o microplástico é persistente no ambiente em níveis altíssimos, particularmente em ambientes aquáticos, pois sua degradação total é muito lenta, podendo levar de centenas a milhares de anos.
Só para termos uma ideia, 35% do microplástico de todo o oceano tem como origem a erosão de poliéster, acrílico ou nylon de roupas. Imagina isso! Podemos refletir um pouco sobre como nos vestimos e qual é o impacto dessas nossas escolhas no ambiente, certo? É simples? Não, claro que não! Sem dúvida, é muito desafiador deixar de usar roupas leves, como camisas UV ou shorts para corrida, e passarmos a usar algodão, não é? Você também pode questionar, “pois é! E onde vão plantar todo esse algodão? Vão terminar de derrubar a Amazônia e o Cerrado?!” Como vocês podem ver, a questão é complexa e não é possível “culpar” simplesmente o consumidor, no caso, você e eu. É importante que pensemos de modo integrado e que possamos vislumbrar uma solução sustentável para este problema, que parece só se acumular.
Tudo bem, já entendemos que o problema é generalizado e fruto do nosso próprio modo de vida, mas qual é o problema de fato de esse “plastiquinho” ficar boiando no mar, depositado no solo e flutuando pelo ar, já que mal conseguimos vê-lo? Por ser muito pequeno, o microplástico tem alta probabilidade de ser ingerido por animais, podendo se acumular em seus tecidos, como ocorre com esponjas, corais, camarões, peixes e até pequenos organismos que vivem no solo. Como outros tipos de poluição, os microplásticos se acumulam na cadeia alimentar, ou seja, um animal que come outro que esteja contaminado também ficará contaminado, estando sujeito a sofrer com seus efeitos tóxicos, que causam, por exemplo, a morte de células. Quando colocamos isso em perspectiva, fica mais clara a importância de procurarmos entender os impactos do microplástico na nossa própria saúde, a curtíssimo prazo.
Voltando ao estudo divulgado pelo The Guardian, os pesquisadores avaliaram a presença de cinco tipos de plástico no sangue de doadores voluntários. Setenta e sete por cento dos pacientes apresentaram microplástico no sangue, sendo que alguns pacientes apresentaram até três tipos de plástico. PET, plástico usado para fabricação de garrafas de água mineral ou refrigerante, por exemplo, foi o tipo de plástico mais prevalente nas amostras. A descoberta de microplásticos em seres humanos não é novidade. Eles já foram encontrados em fezes de adultos e crianças e em praticamente em todos os tecidos humanos estudados, incluindo cérebro e trato gastro-intestinal,. Contudo, eles nunca tinham sido encontrados na corrente sanguínea. Ainda não sabemos ao certo as possíveis consequências para a saúde humana, mas certamente devemos acender um sinal de alerta. Um estudo mostrou que microplástico de PVC (plocloreto de vinila) é mais tóxico para células (linfócitos) de humanos do que de peixes. Alguns efeitos já relatados do microplástico são: deformação de células, morte celular e reações alérgicas, inflamação, distúrbios na microbiota intestinal, desbalanço imunológico entre mãe e feto, causando toxicidade reprodutiva ou danos em neurônios no cérebro.
Já estamos habitando um planeta onde podemos encontrar plástico desde as profundezas do oceano até o cume do Monte Everest, a mais de 8 mil metros de altitude! É estimado que um ser humano consuma em média mais de 50 mil partículas de plástico por ano e, se considerarmos o que é inalado, muito mais. Com tudo isso, e com a projeção de a produção ser o dobro da atual em 2040, torna-se imperativo que o máximo de esforços sejam feitos para que possamos reduzir esse impacto, bem como para que possamos de fato conhecer profundamente o tamanho do problema, o qual ainda não conhecemos suficientemente.
Ao passo em que reconhecemos que o acúmulo de plásticos no ambiente, e agora no corpo humano, é crescente, existem diversas propostas para mitigar o problema ou remediá-lo. Estas propostas incluem filtração dos sistemas de coleta de água, sistemas de coleta automatizada no oceano, reforço na educação e uso consciente de plástico e reciclagem, que constituem algumas das frentes para a possível resolução do problema. Além disso, diversos países vêm tentando reduzir a produção e o consumo de plásticos descartáveis através de leis, como é o caso da China. Há eventos onde a redução da produção e descarte de plástico é praticamente inevitável, precisamos a aprender a lidar com isso. No entanto, situações emergenciais, como a pandemia da Covid-19, podem trazer uma explosão no descarte de produtos plásticos e impor mais um desafio para a redução no consumo de plástico. Um parêntesis: para muitos a pandemia pode parecer uma situação inesperada, mas não é. A comunidade científica previu desde uma década atrás a emergência de uma nova pandemia de origem zoonótica, em virtude das mudanças ambientais causadas pela humanidade. No mesmo ano. foi argumentado que devíamos adaptar estratégias de pesquisa e vigilância para superar desafios relacionados à predição, prevenção e controle de uma futura pandemia. Foi sugerido, além disso, que a futura transmissão direta de coronavírus de morcegos para humanos era um evento provável. A COVID-19 foi a doença que realizou essas predições.
Sem dúvida, um olhar mais consciente sobre o problema que estamos vivendo pode auxiliar a resolvê-lo, mas certamente uma estratégia que seja realmente eficiente necessita incluir o máximo de abordagens possíveis. Por fim, uma abordagem que me parece muito promissora e na qual eu tenho interesse especial é a biodegradação do plástico. Esse processo compreende a ação de microrganismos, como fungos e bactérias decompositores, como é ilustrado no filme “Fungos Fantásticos”. Hoje, possuímos ferramentas e abordagens que nos permitem descobrir organismos que podem ser utilizados para esse propósito, além de termos à disposição técnicas moleculares que nos permitem “engenheirar” organismos já conhecidos para serem ainda mais eficientes.
Pedro Milet Meirelles
Laboratório de Bioinformática e Ecologia Microbiana
Já dizia Paulo Freire “(…) ensinar não é transferir conhecimento, mas criar as possibilidades para a sua própria produção ou construção”. Estudos revelam que a Ciência Cidadã é uma dessas possibilidades, podendo ser usada no ambiente escolar e universitário para promover engajamento com a ciência e atitudes positivas em relação à ciência e ao meio ambiente.
Imagens do Guardiões da Chapada em ação em escolas nos municípios de Lençóis e Mucugê, Bahia (fotos da autora)
É inegável que a ciência está passando por uma crise de confiança nesse começo de século XXI. Os dados da pesquisa realizada pelo Instituto Gallup, em 2019, encomendada pela organização britânica Wellcome Trust, mostram que 75% dos brasileiros entrevistados nessa pesquisa responderam que desconfiam da ciência e 23% consideram que a produção científica pouco contribui para o desenvolvimento econômico e social do país. Quais as explicações possíveis para essa falta de confiança das pessoas na ciência? Especialistas apontam diversas causas, dentre elas, falta de investimentos em uma educação científica crítica desde a educação básica e/ou disseminação de informações falsas, que confundem deliberadamente as pessoas, em nome de interesses escusos, e/ou a atual crise de legitimidade institucional, com as pessoas deixado de acreditar nas instituições republicanas, e a ciência entrando nesse bojo.
As consequências negativas da ausência de valores próprios do pensamento científico são sentidas pelo número cada vez maior de pessoas que acreditam em pseudociências e teorias da conspiração e que se deixam enganar por notícias falsas. A desinformação é ainda mais preocupante quando diz respeito a determinados temas relacionados à saúde pública, como podemos ver, no presente, no aumento da parcela da população que resiste à vacinação. No tocante às questões ambientais, a desinformação também impede que as pessoas enxerguem as relações de interdependência dos seres humanos com o mundo natural. No caso das plantas, por exemplo, a cegueira em relação à forma como elas se reproduzem e sua importância em nossas vidas faz com que as pessoas não se incomodem com a perda da biodiversidade vegetal, provocada por ações humanas, como queimadas e desmatamentos, levando-as a ignorar ou apoiar tomadas de decisão não qualificadas que poderão afetar seriamente as nossas vidas e das gerações futuras.
Em artigo publicado aqui no blog, Charbel N. El-Hani e Tiago Guimarães consideram que a crise de confiança na ciência tem a ver, ao menos em parte, com o modo como ensinamos ciências às pessoas. Na avaliação desses colegas, “se as pessoas entendessem mais sobre o que é a ciência e como a ciência funciona, o conhecimento científico poderia ser usado de forma mais justa e proveitosa”. Eles também sugerem que “não precisaríamos nos preocupar com a desinformação, pois as pessoas filtrariam esses documentos falsos, reconhecendo-os pela falta de marcas evidentes das práticas científicas”. Em outra postagem, El-Hani chama a atenção para o fato de que ensinar como a ciência funciona não consiste em ensinar métodos e fazer experimentos. Para isso, é necessário adotar um conjunto de estratégias e ações que aproximem o estudante das práticas científicas.
A Ciência Cidadã (CC) tem potencial para ser utilizada no ambiente escolar e universitário como uma dessas estratégias. Mas antes de começarmos a discutir como a CC poderia ser incluída como ferramenta na educação científica, é importante saber de qual (ou quais) CC estamos falando, pois esse é um termo cuja definição varia bastante entre as diferentes áreas do conhecimento e regiões geográficas. Aqui estamos adotando uma definição ampla e abrangente, proposta pela Rede Brasileira de Ciência Cidadã, que inclui (…) ”uma gama de tipos de parcerias entre cientistas e interessados em ciência, para produção compartilhada de conhecimentos com potencial para promover: o engajamento do público em diferentes etapas da prática científica; a educação científica e tecnológica, e co-elaboração e implementação de políticas públicas sobre temas de relevância social e ambiental”. Essa definição abarca, assim, uma série de iniciativas ao longo de um espectro entre as duas visões da Ciência Cidadã, a do filósofo e sociólogo inglês Alan Irwin,que define ciência cidadã como “ciência democrática”, e a do zoólogo Rick Bonney, que usa o termo para descrever projetos em que o público se envolve ativamente na investigação científica e na conservação ambiental, o que ele chama de “ciência participativa”.
Dada a pluralidade conceitual, as iniciativas de CC podem ser classificadas de diferentes modos: pelo tipo/nível de colaboração entre cientistas e público interessado em ciência; pela forma de participação na coleta de dados em programas de monitoramentos e inventários da biodiversidade; pelo grau de contribuição do projeto para a ciência; quanto à natureza ou ao tipo de atividade a ser realizada; pelo nível de participação do cidadão voluntário, interessado em ciência, na pesquisa científica; e quanto aos objetivos do projeto de ciência cidadã. Em relação à participação, os projetos contributivos e colaborativos são os mais comuns e, em geral, de maior interesse da comunidade científica, podendo ser acessados em diversas plataformas, como por exemplo iNaturalist, Zooniverse e SibBr. Já os projetos co-criados, embora menos numerosos e em menor escala, possuem maior relevância social.
Mas, independentemente da tipologia, estudos revelam que a participação em projetos de ciência cidadã proporciona aos estudantes um ambiente favorável para o engajamento com a ciência, levando à melhor compreensão da diversidade da pesquisa científica e do pensamento científico, e promovendo atitudes positivas em relação à ciência e ao meio ambiente. Muito embora os projetos colaborativos contribuam mais para aprendizagem de conteúdos específicos e para aumentar o interesse nesses conteúdos, os projetos co-criados contribuem mais para o desenvolvimento de habilidades para a investigação científica.
No escopo do INCT IN-TREE, alguns projetos de CC estão sendo conduzidos, dentre eles, os projetos Guardiões da Chapada e Guardiões dos Sertões de Sergipe. Em linhas gerais, os “Guardiões” são projetos colaborativos de ciência cidadã que envolvem voluntários no monitoramento participativo da interação planta-visitante em ambientes naturais, urbanos e agrícolas, por meio do registro fotográfico das interações.
Apesar das especificidades de cada projeto “Guardiões”, no que diz respeito aos locais de atuação, aos tipos de ambientes monitorados e às ações que desenvolvem, eles têm em comum o fato de estarem baseados em três pilares, que coexistem e interagem entre si de forma harmoniosa, organizados em duas dimensões, educacional e científica, quais sejam: 1) Produção colaborativa de informações de base científica, por meio do monitoramento participativo, que permite gerar novos conhecimentos sobre a biodiversidade; 2) Tradução e compartilhamento dos conhecimentos relacionados aos projetos ou gerados por estes; 3) Despertar da cidadania científica e ambiental, por meio de ações voltadas à capacitação, ao desenvolvimento de habilidades e ao engajamento em ações de conservação.
Na dimensão científica do projeto, a colaboração entre cientistas e interessados em ciência envolve a coleta de dados das interações por meio do registro fotográfico. As fotos podem ser tiradas com uma câmera fotográfica, com o celular ou com o tablete e enviadas diretamente para uma plataforma online E-Guardiões da biodiversidade, ou por meio do aplicativo “Guardiões da Biodiversidade”, desenvolvido para dispositivos móveis, já disponíveis para IOS e Android, o qual permite também registrar as interações em tempo real. As fotos tiradas, com as respectivas informações, são armazenadas em um banco de dados, onde especialistas poderão acessá-las e fazer a identificação dos animais e de plantas e complementar a descrição da interação planta-visitante, se for necessário. Com todos esses dados, será possível, por exemplo, mapear novas ocorrências de espécies, avaliar o efeito das mudanças ambientais sobre as espécies, desenvolver diretrizes para manejo e conservação da vida silvestre, e planejar o uso sustentável da terra na região. Como um projeto científico, os projetos “Guardiões” devem produzir dados e informações científicas genuínas, que possam gerar novos conhecimentos. Nesse sentido, esses projetos buscam aliar relevância e rigor por intermédio da parceria entre cientistas profissionais e demais atores sociais. A parceria com cientistas ajuda a aumentar a confiança e a credibilidade nos dados, enquanto a parceria com os atores sociais aumenta a relevância e o impacto social dos resultados.
No ambiente escolar e universitário, a dimensão científica dos projetos “Guardiões” pode ser explorada para colocar os estudantes em contato com a prática científica e a natureza da ciência, estimulando o estudante a formular questões situadas na sua realidade local e a discutir os resultados das pesquisas. As questões podem variar conforme o perfil e nível de escolaridade dos estudantes, o que fica a cargo do professor, que deve fazer as adequações que julgar necessárias.
Na dimensão educacional, os temas centrais dos projetos Guardiões podem ser contextualizados por situações ou problemas socioambientais, locais ou regionais, como, por exemplo, desmatamento; queimadas; uso de agrotóxicos; produção de alimentos; pragas e doenças de cultivos agrícolas; declínio de polinizadores; qualidade do ar e da água; arborização em espaços urbanos e bem-estar humano; mudanças climáticas e a biodiversidade; espécies invasoras, dentre outros. Dessa maneira, podem contribuir para tornar os conhecimentos escolares mais significativos para os estudantes. Essas situações-problemas poderão mobilizar tanto conhecimentos científicos quanto outros tipos de conhecimentos, necessários para busca de soluções, o que favorece uma aprendizagem multidimensional dos conteúdos, que permite ao estudante, além de compreender conceitos e princípios técnicos e memorizar fatos e evidências, também exercitar procedimentos e ações práticas e vivenciar e refletir sobre situações concretas, do mundo real.
Ao relacionar os conceitos científicos aprendidos em sala de aula às questões científicas relevantes para a vida das pessoas, ou sejam, às questões sociocientíficas, os projetos Guardiões também contribuem para aproximar a escola/universidade do seu entorno, promover uma educação científica crítica dos estudantes, desenvolver novas habilidades e competências, despertar novos talentos para a carreira científica, e construir os valores necessários para tomada de decisões responsáveis sobre questões relacionadas às relações entre ciência e tecnologia na sociedade e no ambiente (CTSA).
Contudo, existem desafios para implementar projetos de CC nas escolas, dentre eles, a necessidade de capacitação dos professores, bem como dos cientistas, para co-produção e condução dos projetos; adaptação do projeto aos currículos e processos avaliativos; dificuldades para garantir a qualidade dos dados coletados pelos estudantes, bem como para análise e interpretações dos resultados; e falta de recursos financeiros contínuos para garantir a sustentabilidade do projeto e o tempo de professores e pesquisadores para dedicação aos projetos. Por sua vez, esses desafios podem ser em parte solucionados pelo estabelecimento de parcerias entre Universidade-Escola, entre professores de diferentes disciplinas e, se possível, entre as escolas ou universidades e diferentes setores da sociedade; pela formulação de objetivos científicos e educacionais claros e protocolos simples, adequados ao perfil e nível de escolaridade dos estudantes; por ações que incentivem a participação dos estudantes e dos professores desde o início do projeto; pela transparência e compartilhamento dos resultados; e por atividades que visem manter engajamento, interesse e motivação de todas as partes interessadas no projeto.
Buscando contribuir para superação desses desafios, os projetos “Guardiões” vêm construindo parcerias com instituições de ensino, organizações não-governamentais e coletivos para o desenvolvimento de ações educacionais. Na Bahia, o Guardiões da Chapada tem atuado nos municípios de Lençóis, Mucugê, Ibiocoara e Piatã no âmbito de uma Ação Curricular em Comunidade e Sociedade (ACCS), componente curricular, na modalidade disciplina, que visa ampliar a possibilidade e a qualidade de participação pública em temas relacionados ao uso e à conservação da biodiversidade e dos serviços ecossistêmicos, instrumentalizando os atores sociais para intervirem melhor nos processos decisórios relacionados à conservação ambiental; contribuir para a formação de profissionais comprometidos e engajados com a solução dos problemas ambientais; e estimular a autonomia e criticidade dos participantes, na busca de soluções sistemáticas para problemas socioambientais. Em Sergipe, as ações do Guardiões dos Sertões acontecem por intermédio de parcerias construídas pelo projeto “Protagonistas da Escola Verde: experiência de aprendizagem em ecologia e educação científica no ensino médio”, que articula a Universidade Federal de Sergipe, escolas da educação básica, gestão pública e comunidade em geral em prol de avanços na discussão de temas relacionados à ética, à biodiversidade e aos serviços ecossistêmicos, no contexto do semiárido sergipano, com abordagem participativa. Interessados em conhecer essas ações podem visitar as nossas páginas nas redes sociais e/ou entrar em contato com os coordenadores dos projetos “Guardiões” através do e-mail guardioeschapada@gmail.com
Viana, BF; Souza, CQ. Ciência cidadã para além da coleta de dados. Revista ComCiência – Revista eletrônica de jornalismo cientifico, Labjor, UNICAMP, p. 1 – 2, 05 out. 2020.
As inversões cromossômicas são de grande interesse na biologia pois estão comumente associadas à origem de inúmeros fenômenos, como organização social, adaptação ambiental, isolamento reprodutivo e até especiação. Duas características fazem delas instrumentos eficazes para adaptação local: envolvem muitos genes e reduzem drasticamente a permutação quando estão em heterozigose. Juntas, essas características produzem um cenário favorável para a disseminação de genes envolvidos na adaptação local em populações naturais.
Por volta de 1915, na famosa sala das moscas da Columbia University, pesquisadores perceberam que alguns estoques de Drosophila melanogaster apresentavam uma taxa de permutação (“crossing-over”) muito menor que a esperada. Alfred Sturtevant propôs que havia fatores supressores da permutação e, por meio de cruzamentos e contagem de mutantes de D. melanogaster, observou que a ordem dos genes nos estoques com diferentes taxas de permutação também era diferente. Essa observação o levou a concluir que os fatores inibidores de permutação eram inversões de regiões dos cromossomos. Foi então levantada uma hipótese para explicar essa observação: a permutação era impedida pois os cromossomos homólogos não poderiam se emparelhar na região invertida durante a meiose. A hipótese foi logo descartada após a observação do emparelhamento (em forma de alça, figura 1) em milho e em cromossomos politênicos de Drosophila. O emparelhamento ocorre e a permutação também ocorre, mas os produtos recombinantes tornam os gametas inviáveis na grande maioria das vezes e, por isso, não são observados indivíduos gerados por esses gametas (figura 1).
Figura 1. Esquema mostrando os dois tipos de inversão cromossômica, ambas em heterozigose, e os gametas resultantes da permutação. (A) a porção cromossômica invertida pode não incluir o centrômero (inversão paracentromérica). Neste caso, a permutação dá origem a uma cromátide sem centrômero e outra com dois centrômeros. Cromátide sem centrômero não fica ligada às fibras do fuso e não é puxada para os pólos da célula durante a divisão meiótica. Por outro lado, a cromátide com dois centrômeros é puxada para os dois pólos e se rompe. Essas cromátides recombinantes não geram gametas viáveis. (B) Se a inversão inclui o centrômero (inversão pericentromérica), a permutação leva a criação de cromátides recombinantes que ou não possuem todos os genes, ou cromátides com genes duplicados, que por sua vez não formam gametas viáveis. Reproduzido de Moore JA (1986) Science as a Way of Knowing – Genetics. American Zoologist, McLean, 26, 583-747 [texto traduzido e adaptado pelos docentes da disciplina Genética (E.J.C. de Almeida; J.M. Amabis; M.L. Benozzati; B.C. Bitner-Mathé; E.M. Dessen; C.F.M. Menck; L. Mori; C.R. Vilela & Y.Y. Yassuda), do IB-USP, em 1995; revisado em 2021].
A supressão da permutação durante a meiose (ou melhor, a supressão da produção de gametas recombinantes) e, consequentemente, do rearranjo de alelos, pode dar origem a supergenes que “prendem” alelos de diferentes genes juntos em grandes regiões cromossômicas. Quando as inversões capturam alelos de diferentes genes que conferem maior aptidão, essas variantes invertidas rapidamente se tornam mais frequentes nas populações. Muitas delas explicam o surgimento de diversos fenótipos, como, por exemplo: diferenças reprodutivas e de comportamento em machos da ave combatente, Philomachus pugnax; coloração e comportamento no pardal-de-garganta-branca, Zonotrichia albicollis; ciclo de vida, morfologia e tempo de floração em Mimulus guttatus; ecologia e morfologia do peixinho esgana-gata, Gasterosteus aculeatus; formas de organização social na formiga-de-fogo, Solenopsis invicta; e resistência a inseticidas em Anopheles arabiensis. Um exemplo de como as inversões facilitaram a disseminação de alelos de resistência a inseticidas foi descrito em mosquitos do gênero Culex. Os alelos que conferem resistência são deletérios quando estão em homozigose e adaptativos em heterozigose. Esses alelos podem ser potencialmente mantidos em um estado heterozigótico por meio da presença de inversões. Muitas dessas inversões segregam dentro das espécies por centenas de milhares ou mesmo milhões de gerações. Por exemplo, uma inversão de 900 kb no cromossomo 17q21.31 de humanos é observada em europeus e alguns asiáticos (haplótipo H2). Os dois haplótipos, H1 e H2, que divergiram há três milhões de anos, são anteriores ao surgimento do homem moderno e até mesmo à origem do gênero Homo. A explicação mais empregada para a longa retenção de polimorfismos de inversão é a seleção balanceadora.
A espécie de borboleta Heliconius numata tem um dos exemplos mais fascinantes de variação fenotípica associada a inversões. A espécie apresenta sete padrões de asa, mimetizando sete espécies do gênero Melinaea, outro grupo de borboletas, mais tóxicas para seus predadores (figura 2). As sete diferentes formas de H. numata podem aparecer um uma única população. Essas diferentes formas são resultado de vários rearranjos no cromossomo 15. A inversão inicial, denominada supergene P, oferece vantagens por meio do mimetismo e provavelmente entrou na população por meio de hibridação com a espécie Heliconius pardalinus. Após o rearranjo inicial, a região cromossômica não permaneceu estática. Outras duas inversões ocorreram em regiões adjacentes ao supergene P, dando origem a novos haplótipos (alelos de locos adjacentes que são herdados como uma unidade) e novos padrões de asa. A arquitetura do supergene é caracterizada por um bloco não recombinante que captura 21 genes distintos envolvidos no padrão de asa, que são conhecidos por se recombinarem em outras espécies do grupo. Nas espécies relacionadas H. melpomene e H. erato, há vários locos independentes (e em cromossomos distintos) envolvidos no padrão de asa.
Figura 2. Cada uma das formas polimórficas de H. numata mimetiza espécies diferentes do gênero Melinaea (painel superior). Cada forma é controlada por um alelo do supergene P, com dominância crescente mostrada da esquerda para a direita (painel central). Em todas as outras espécies estudadas no gênero Heliconius, o padrão das asas é controlado por vários locos de grande efeito em diferentes cromossomos, a exemplo de H. melpomene, mostrada no painel inferior. Figura reproduzida de Joron et al. (2011).
Mesmo conferindo uma grande vantagem seletiva, o supergene não foi fixado nas populações de H. numata após sua origem e pesquisadores mostraram no inicio deste ano por que isso acontece. O haplótipo seletivamente favorecido (com os alelos adaptativos dos diferentes genes ligados) se espalha pela população, mas, devido à falta de permutação, acumula uma série de mutações com padrão de herança recessiva que diminui a aptidão dos homozigotos (a relação entre a permutação e o acúmulo de mutações foi explorada em um post anterior, aqui no Darwinianas). Dentre as principais mutações observadas pelos pesquisadores, está o acúmulo gradual de elementos transponíveis de diferentes classes. Como consequência, os haplótipos não conseguem atingir a fixação, mas persistem na população, predominantemente em indivíduos heterozigotos.
Para testar o efeito das inversões nas borboletas, os autores analisaram a sobrevivência de larvas de cruzamentos de tipos diferentes sob condições controladas de laboratório. Eles observaram que a sobrevivência de larvas homozigotas para os rearranjos era muito reduzida: apenas 6,2% das larvas homozigota para uma das inversões secundárias e 31,3% homozigotas para a outra inversão secundária sobreviveram até o estágio adulto. Larvas heterozigotas com dois haplótipos diferentes mostraram uma recuperação na aptidão e sua sobrevivência era quase indistinguível daquela de larvas homozigotas para o arranjo ancestral (77,6% de sobrevivência). As inversões, portanto, abrigam variantes recessivas com um forte impacto na sobrevivência individual em homozigotos. Essa descoberta implica que os diferentes haplótipos invertidos não compartilham as mesmas mutações deletérias, mas, em vez disso, rearranjos subsequentes podem compensar as mutações que se acumularam ao longo do tempo. Consequentemente, novos rearranjos (ou eventos raros de recombinação dentro da região invertida) podem estender a vida do haplótipo. A baixa aptidão larval dos homozigotos também sugere que o potencial de qualquer um desses haplótipos se tornarem fixos é bastante baixo. Assim, a seleção é dependente da frequência, beneficiando haplótipos presentes em menor frequência. Como consequência, esse mecanismo garante que as diferentes inversões coexistam ao longo do tempo e também deve estar envolvido na manutenção de supergenes observados em outras espécies, como o pardal-de-garganta-branca e a formiga-de-fogo, mesmo com alta letalidade nos homozigotos.
Outra revisão, também em inglês, os autores trazem estudos recentes em genômica das inversões polimórficas em animais e plantas para detalhar as causas e consequências da persistência de inversões polimórficas na natureza.