
Como já dizia o ditado, o que os olhos não vêem, o coração não sente. Talvez por isso o “mundo subterrâneo” seja ainda tão pouco estudado. Mas, a diversidade de vida “escondida” no solo é impressionante: ela representa aproximadamente ¼ de toda a vida do planeta, e sabemos ainda muito pouco sobre ela. Conhecemos apenas algo em torno de 1% dos microorganismos presentes no solo. No entanto, há mais de duas décadas, sabemos que a saúde dos solos e da biodiversidade ali presente é fundamental para o funcionamento adequado dos ecossistemas terrestres (veja aqui um outro post do Darwinianas onde discuto a importância dos solos para os ecossistemas terrestres). A biodiversidade do solo participa de processos essenciais como a decomposição de matéria orgânica e a ciclagem de nutrientes, a fertilidade e a disponibilidade de água.
Em particular, a fração microscópica desse mundo subterrâneo tem grande importância para a sobrevivência de muitas espécies de plantas. Por exemplo, a maior parte do nitrogênio atmosférico fixado em ambientes terrestres provém da associação simbiótica entre bactérias e plantas. Bactérias simbiontes, dos gêneros Rhizobium, Bradyrhizobium, Frankia, e Azospirillum, são exemplos das chamadas bactérias fixadoras de nitrogênio e realizam essa função por meio de simbiose com raízes de plantas no solo.
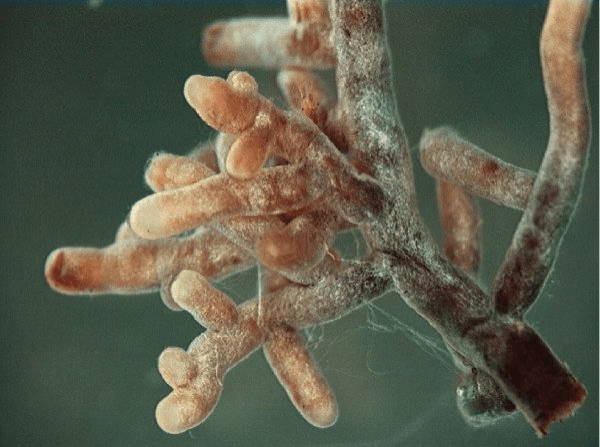
Um outro grupo de organismos microscópicos merece atenção especial: os fungos capazes de formar associações simbióticas com raízes de plantas chamadas de micorrizas. Essas associações podem ser de dois tipos principais: as ectomicorrizas e as endomicorrizas. Enquanto nas associações do tipo ectomicorriza os fungos, geralmente dos filos Basidiomycota e Ascomycota, se alojam entre as células da planta hospedeira, nas associações endomicorrizas espécies de fungos, principalmente do filo Glomeromycota, são capazes de penetrar a célula vegetal.
Ectomicorrizas ocorrem em associação a aproximadamente 2% das espécies de plantas, e participam da captação de água e nutrientes, geralmente ajudando a sobrevivência das plantas em condições pouco favoráveis. Já as endomicorrizas, também chamadas de micorrizas arbusculares, ocorrem em 80% das famílias de plantas e estão entre as associações simbióticas mais prevalentes do planta. Essa associação auxilia a captação de fósforo, enxofre, nitrogênio e micronutrientes pela planta hospedeira.
Essa semana, um grupo de pesquisadores produziu um mapa global da distribuição de simbiontes de espécies florestais, com base em um banco de dados de mais de 1 milhão de amostras de florestas, e incluindo mais de 28 mil espécies arbóreas. Os pesquisadores estimaram que árvores associadas a ectomicorrizas, apesar de representarem apenas 2% de todas as espécies de plantas conhecidas, constituem aproximadamente 60% das árvores do planeta. Simbiose do tipo ectomicorriza domina florestas nas quais o clima frio e seco inibe a decomposição e é a principal forma de simbiose em alta latitude e grandes elevações. Em contraste, as espécies associadas a micorrizas arbusculares dominam florestas tropicais úmidas, e ocorrem em associação a ectomicorrizas apenas em biomas temperados, nos quais a sazonalidade (com períodos de clima quente e úmido) aumenta as taxas de decomposição. Já as bactérias fixadoras de nitrogênio, usualmente insensíveis ao controle climático nas taxas de decomposição, são mais abundantes em zonas áridas com solos alcalinos e altas temperaturas (Figura 1).

O modelo criado pelos pesquisadores prediz também transições abruptas entre os tipos dominantes de simbiontes, largamente associadas às taxas de decomposição controladas pelo clima. Essas transições, sugerem os pesquisadores, são provavelmente causadas por retroalimentação positiva entre as plantas e os microorganismos do solo. Assim, as taxas de decomposição dependentes do clima são a principal força motriz determinando a distribuição de simbiontes de espécies de árvores no globo (Figura 2).

Os dados apresentados por Steindinger e colaboradores são consistentes com a chamada hipótese de Read, que propõe que as baixas taxas de decomposição em altas latitudes e elevações favorecem o estabelecimento de ectomicorrizas devido à sua capacidade de liberação de nutrientes orgânicos no solo. A hipótese de Read foi a primeira proposta da existência de uma correlação entre os ciclos biogeoquímicos e os microorganismos simbiontes. Talvez também por sua importância histórica, os autores propõem que o gradiente de elevação e latitude associado à transição de florestas dominadas por endomicorrizas para florestas dominadas por ectomicorrizas seja chamado de Regra de Read.
Os autores apontam ainda para a extensão global do chamado “padaroxo da ciclagem de nitrogênio”. Esse paradoxo refere-se ao fato de que, apesar de o nitrogênio ser um fator limitante mais significativo em zonas temperadas, espécies arbóreas associadas a fixadores de nitrogênio são mais prevalentes em ambientes tropicais. Associações com fixadores de nitrogênio são particularmente comuns em florestas com temperaturas máximas anuais acima de 35°C e solos alcalinos, como em regiões desérticas (veja Figura 1, Arbustos de deserto). O modelo apresentado pelos autores prevê um aumento de duas vezes no número de fixadores de nitrogênio na transição entre ambientes úmidos e florestas tropicais secas, reforçando a ideia de que os fixadores de nitrogênio são importantes simbiontes em ambientes áridos. Os dados não resolvem o paradoxo, e não são capazes de testar as diferentes hipóteses alternativas, capazes de explicar esse fenômeno. Mas, os dados suportam claramente a ideia de que esse paradoxo é um fenômeno global.
As sínteses de tamanha quantidade de dados nos ajudam não apenas a testar ideias antigas, como as descritas acima. Essas sínteses são essenciais para que possamos finalmente “enxergar” os padrões globais de distribuição dos microrganismos. Nesse caso em particular, a compreensão dos padrões globais de simbiontes de espécies arbóreas, e dos principais determinantes dessa distribuição, pode nos ajudar a entender, por exemplo, quão facilmente as árvores podem acessar nutrientes, sequestrar carbono, ou suportar os efeitos das mudanças climáticas. Essa associações simbiontes, muitas vezes invisíveis, podem guardar o futuro das nossas florestas.
Ana Almeida
(California State University East Bay, CSUEB)
Para saber mais:
Bardgett, R. D. 2002. Causes and consequences of biological diversity in soil. Zoology, 105(4): 367-375.
Brundrett, M.C.; Tedersoo, L. 2018. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytologist, 220: 1108–1115.
The factory of life: why soil biodiversity is so important. 2010. European Union.
Imagem: Ectomicorriza do tipo Amanita nas pontas das raízes de uma planta. Fonte: Ellen Larsson, Wikipedia.

.jpg){kind=link}
Uma consideração sobre “Simbiontes, ávores, decomposicão e mudanças climáticas: Associações invisíveis e o futuro das florestas”