

Ao observar atentamente a variação na forma dos animais e plantas na natureza, ou em uma visita a um renomado museu de ciências naturais, imediatamente chama a atenção –não importando se o observante é leigo ou cientista – o quão adequado é o ajuste dos organismos ao seu ambiente.
Até meados do século XIX a explicação para esse “design perfeito” evocava uma divindade. Mas em 1859 o naturalista inglês Charles Darwin mudou essencialmente a forma como pensávamos, propondo que a diversidade observada poderia ser explicada em grande parte pela combinação de mutações aleatórias (variabilidade genética) e a seleção não aleatória dessa variação pelo processo de seleção natural. E isso valia para nós humanos também!

Hoje pode parecer simples essa lógica, mas em 1859 a fonte de variação (ou seja, as mutações) e, mais precisamente, o mecanismo pelo qual essa variação era passada através das gerações, era absolutamente desconhecido. Embora o famoso trabalho de Gregor Mendel que mostrava padrões de herança simples em ervilhas lisas e enrugadas tivesse sido publicado em 1866, passaram-se quase 40 anos até que fosse redescoberto ou plenamente considerado. Ou seja, não havia noção da existência de cromossomos, genes ou mesmo o DNA. Mas Darwin notou que os descendentes se pareciam com seus parentais, portanto, ele sabia que os traços podiam ser transmitidos entre gerações; só não tinha ideia de como isso acontecia. Quase cem anos depois, o link que faltava, ou seja, a unidade fundamental de informação hereditária e fonte de novas variações, foi revelado por Francis Crick e James Watson (que descobriram em 1953 a estrutura tridimensional da dupla hélice, demonstrando que o DNA poderia se replicar).
Desde então a ciência se moveu muito adiante e acumulou grandes conhecimentos na área da genética. Entretanto, as mesmas questões que motivaram Darwin continuam inspirando gerações de cientistas: como e por que os organismos têm morfologias e comportamentos tão distintos? A grande diferença é que hoje conseguimos buscar as respostas a essas questões no DNA, revelando mudanças precisas que permitem aos organismos se adaptarem ao ambiente (Figura 1).
Já em 1900 os fundadores da genética de populações debatiam, através de modelos matemáticos e estatísticos como a adaptação ocorre: se através de grandes saltos (ou seja, um pequeno número de mutações, cada uma com efeito grande no fenótipo) ou pequenos passos (isto é, acumulo de diversas mudanças de menor efeito). Esse tema ainda é amplamente discutido na ciência, que busca também esclarecer se a adaptação procede através de mutações recessivas ou dominantes; se os genes envolvidos agem independentemente ou interagem para produzir características adaptativas, se mutações benéficas tendem a afetar a função da proteína (ou seja, mutações na própria proteína) ou sua expressão espacial ou temporal (ou seja, mutações fora da proteína que controla sua regulação).
Se o cuidadoso observante tiver a oportunidade de ver diversos indivíduos de uma mesma espécie lado a lado, como na coleção científica de um museu de história natural, muito provavelmente notará que existem variações morfológicas também dentro das espécies, e ainda traços muito similares em espécies distantes evolutivamente. Vamos pensar que se um dado fenótipo ocorre repetidas vezes na natureza (de forma independente), não seria de se perguntar com que frequência à seleção natural depende dos mesmos genes e/ou mutações para conduzir a traços similares em diferentes populações ou espécies?
Até meados de 1990 se considerava que as grandes diferenças fenotípicas eram resultado de inúmeras substituições gênicas, cada uma com efeito relativamente pequeno. No entanto, a partir da possibilidade de sequenciamento de DNA em larga escala os estudos com genes que regulam o processo de desenvolvimento, e codificantes de receptores e sinalizadores de complexas redes metabólicas, sugerem que poucas mutações podem ter grande efeito, afetando diretamente a morfologia.
A viagem a bordo do Beagle pela América do Sul e Central em 1831 proporcionou um laboratório vivo para o Darwin. Dentre tantos exemplos de diversidade fenotípica um em especial chamou a atenção do naturalista: a variação na forma e tamanho do bico dos tentilhões nas distintas ilhas do arquipélago de Galápagos que notoriamente estava relacionado a uma variedade de tipos de alimentação (Figura 2). Mais adiante os tentilhões de Darwin se tornaram um exemplo clássico de diversificação de espécies através da seleção natural.

Curiosamente, nos últimos 15 anos, ferramentas moleculares modernas permitiram determinar o papel dos genes Bmp4, Calmodulina e ALX1 na variação craniofacial dos tentilhões de Darwin, cujas diferenças no padrão de expressão dos genes ao longo do desenvolvimento foram associadas à forma e tamanho do bico. Esse é um exemplo clássico de bem sucedida associação causal de um dado fenótipo morfológico e um genótipo específico.
Ainda na rica região Neotropical encontramos outro exemplo de variação morfológica associada à dieta: os morcegos da família Phyllostomidae. Também conhecidos como filostomídeos, representam um dos grupos mais diversos não só entre os morcegos, como mamíferos em geral. O mais curioso são as especializações desse grupo, que se alimentam desde néctar, frutas, insetos, carne, grãos até sangue. Uma análise até mesmo superficial da estrutura craniofacial permite perceber alterações na mandíbula e palato (e também dentição) específicas para cada tipo alimentar, também incluindo tamanho e forma como no bico dos tentilhões de Galápagos (Figura 3). A mudança mais caraterísticas é o focinho alongado nas espécies que se alimentam de néctar. E será que as alterações morfológicas craniofaciais nos morcegos poderiam ter a mesma base genética identificada nos tentilhões?

Poderíamos, portanto, pensar no papel dos mesmos genes, pois seriam potenciais candidatos a explicar tal variação. Mas com o avanço de estudos em modelos animais e de genética humana clínica, emergiram outros genes do desenvolvimento específicos de alterações na morfologia craniofacial que são melhores candidatos a explicar a origem da variação observada nesse grupo de morcegos, por exemplo, os genes do fator de crescimento fibroblástico (FGF), e fatores de transcrição das famílias RUNX e PAX.
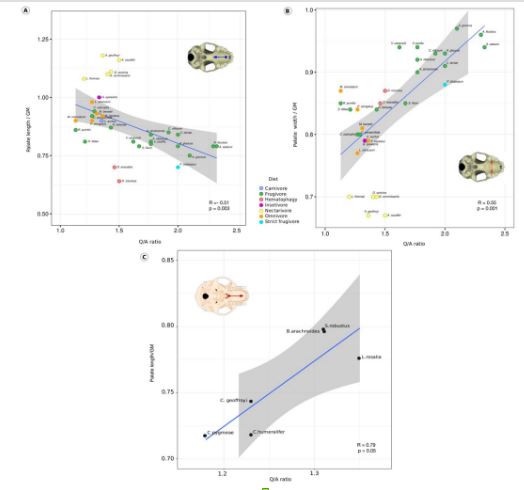
Vamos considerar especificamente no gene RUNX2 (fator de transcrição relacionado ao Runt 2), que já foi demonstrado ter grande efeito na variabilidade craniofacial dos mamíferos, associado à formação do tecido ósseo, portanto, essencial para a osteogênese. O RUNX2 contém uma região de repetição de aminoácidos em tandem que precede o domínio, com um segmento rico em glutamato (poli Q), seguido por outro rico em alanina (poli A) que desempenha papel importante na ativação e repressão de proteínas essenciais para o desenvolvimento craniofacial. Embora estudos prévios com cães de raça e carnívoros silvestres de distintas morfologias tenham encontrado uma correlação positiva entre a proporção de Q para A no domínio de repetição desse gene (também chamada de taxa Q/A) e o comprimento craniofacial, quando se analisou mamíferos placentários não carnívoros e marsupiais, não foi verificado tal correlação.
Voltando a incrivelmente diversa linhagem de morcegos filostomídeos. A variação morfológica craniofacial é significativa não somente entre grupos de espécies que formam guildas alimentares, mas também dentro de alguns desses grupos, como aquele dos frugívoros. E essa admirável variação também foi recuperada na taxa Q/A do RUNX2 dos morcegos. A diferença é que pela primeira vez foi demonstrada a associação desse gene com o encurtamento e alargamento do rosto dos filostomídeos (Figura 4), ou seja, o oposto do padrão (=aumento no comprimento) verificado em cães, carnívoros e também em primatas do novo mundo.
Esse exemplo nos mostra que a conexão de uma dada característica morfológica com um genótipo causal é mais complexa ainda. Notamos, portanto, que em determinados casos não se trata de encontrar as mesmas mutações (ou padrões de expressão diferencial) em espécies distintas, mas sim um ajuste fino no mesmo mecanismo (como o gene RUNX2) para explicar as bases de fenótipos similares. Portanto, o processo adaptativo pode usar de formas alternativas de um mesmo caminho, ou mesmo caminhos distintos, para chegar ao mesmo lugar.
A ciência tem se movido a passos largos desde os achados de Darwin e Watson e Crick. Mas para cada pergunta respondida desdobram-se novas dúvidas. Enquanto relevamos as bases moleculares do que Darwin chamava de “aquela perfeição de estrutura e co-adaptação que justamente estimula nossa admiração”, aprendemos com a espetacular evolução da diversidade morfológica dentro e entre as espécies.
Gislene Lopes Gonçalves
UFRGS e Universidad de Tarapacá, Chile
Sugestões de leitura:
- Ferraz T, Rossoni DM, Althof SL, Pissinatti A, Paixão Cortês VR, Bortolini MC, González-José R, Marroig G, Salzano FM, Gonçalves GL, Hünemeier T (2018). Contrasting patterns of RUNX2 repeat variations are associated with palate shape in phyllostomid bats and New World primates. Scientific Reports 8:7867
- Hoekstra H (2010). From Darwin to DNA: The Genetic Basis of Color Adaptations. In: In the Light of Evolution: Essays from the Laboratory and Field (Ed. Losos J). Roberts and Company Publishers.
- Lamichhaney S, Berglund J, Almén MS, Maqbool K, Grabherr M, Martinez-Barrio A, Promerová M, Rubin C-J, Wang C, Zamani N, Grant BR, Grant PR, Webster MT, Andersson L (2015). Evolution of Darwin’s finches and their beaks revealed by genome sequencing. Nature, 518: 371–375.
