
Sem a evolução da multicelularidade e, mais especificamente, dos animais, claro que eu não estaria aqui escrevendo este texto. Nós não estaríamos aqui. Uma interpretação evolutiva das origens (dos organismos multicelulares, dos animais, dos humanos etc.) sempre causa controvérsia, mas pode haver pouca dúvida de que ela nos mostra como podemos ser (e somos) bem sucedidos em explicar questões tão fundamentais quanto a das origens de uma perspectiva naturalista – isto é, que não admite quaisquer outras causas que não causas naturais.
Desta perspectiva, não somos consequências necessárias da história da Terra, quanto mais do universo. Somos produtos contingentes dessa história. Existimos, mas poderíamos sem dúvida não existir. E nada haveria de errado na ordem das coisas. Longe de desencantar os mistérios de nossa existência, esta é uma visão que pode nos encher de espanto. É de fato espantoso que estejamos aqui, tantas as possibilidades de não estarmos: se a vida não tivesse surgido no planeta, se estivesse sido extinta antes de nosso surgimento, se os eucariotos não tivessem evoluído (com suas células dotadas de sistemas de membranas internas), se os animais, se os cordados, se os primatas não tivessem evoluído, nós não estaríamos aqui.
Assim, o surgimento dos animais, que poderia parecer apenas um evento distante, de pouca relevância para nossas vidas, se reveste de suma importância. Ele é parte da explicação de onde viemos. Parte da explicação de por que você está aí, agora, lendo esse texto.
De onde vieram os animais? Esta é uma questão que tem nos desafiado há muito tempo, mas sobre a qual temos cada vez mais conhecimento e boas hipóteses. Uma parcela importante destas hipóteses surgiu da pena do biólogo evolutivo de Oxford Thomas Cavalier-Smith. Em seu artigo mais recente, ele ergue a voz contra o que denomina uma “falta de síntese imaginativa mas crítica, como foi feito por Darwin e Simpson, frequentemente negligenciada de modo prejudicial como especulação e dissuadida por práticas de arbitragem e publicação em periódicos”. Não poderia concordar mais: a ciência anda mesmo pouco imaginativa e demasiado presa a uma mera recitação de dados. Mas não é este o sentimento depois de ler um artigo de Cavalier-Smith. Ali não há escassez de hipóteses ousadas mas plausíveis, por estarem assentadas em profundo conhecimento teórico e empírico. Decerto parte delas não se sustentarão diante das evidências, mas é para isso mesmo que devem ser propostas: para serem testadas e, caso se sustentem, abrir novos horizontes ao pensamento e à imaginação, e, caso não se sustentem, informar nossa busca de novas hipóteses. Antes ter ideias ousadas que podem ser falsas do que não ter mais do que seguras ideias triviais.
Aos 74 anos de idade, Cavalier-Smith oferece, em Origin of animal multicellularity: precursors, causes, consequences – the choanoflagellate/sponge transition, neurogenesis and the Cambrian explosion (Origem da multicelularidade animal: precursores, causas, consequências – a transição coanoflagelado/esponja, neurogênese e a explosão do Cambriano), uma espécie de síntese de várias hipóteses sobre a evolução dos animais que propôs ao longo de sua carreira. São muitas ideias, como o longo título do artigo sugere, e não há como fazer justiça a todas elas aqui. Desse modo, esta postagem fará um recorte no muito que Cavalier-Smith tem a dizer. Recomendo ao leitor ávido por novas e interessantes ideias sobre a evolução animal a se debruçar sobre o artigo original, publicado em Philosophical Transactions of the Royal Society no mês passado.
Como nos lembra Cavalier-Smith, há muito mais unicelulares do que multicelulares na Terra, seja em quantidade, seja em diversidade, cumprindo papeis de importância central na biosfera, por exemplo, nos ciclos biogeoquímicos. Há razões ecológicas e evolutivas para isso: há muitos nichos ecológicos excelentes para unicelulares, enquanto a multicelularidade é frequentemente desvantajosa, como mostram muitos casos em que linhagens unicelulares evoluíram a partir da condição multicelular (por exemplo, múltiplas vezes nas leveduras). Entender como e por que a multicelularidade evoluiu, não uma mas algumas vezes na história da vida, requer a proposição de fortes vantagens seletivas, que contrabalancem as desvantagens da vida multicelular.
Este é um ponto central dos argumentos de Cavalier-Smith: a evolução da multicelularidade é, em termos de sua origem, muito simples: todos os grupos de organismos unicelulares têm um potencial celular e mutacional para se tornar multicelular, por meio de duas vias possíveis: em células que não têm parede celular, como as dos animais e dos bolores limosos (slime molds), evoluem moléculas que as grudam umas nas outras; em células com parede celular (como as das plantas), evoluem modificações da biogênese da parede que inibem a separação das células filhas. O problema não é exatamente de origem, mas de manutenção: quais vantagens seletivas poderiam fixar a multicelularidade numa linhagem originalmente unicelular?
O aspecto central a considerar – argumenta Cavalier-Smith – é o modo de alimentação. Se o organismo for fototrófico (usando fótons, ou seja, luz para adquirir energia para suas funções vitais, por exemplo, mas não exclusivamente, através de fotossíntese), ou se for osmotrófico (nutrindo-se pela absorção de compostos orgânicos por osmose, como fazem muitos fungos), não há dificuldade em tornar-se multicelular: essencialmente a alimentação prossegue do mesmo modo como na unicelularidade. Uma alga ou um fungo constituído por filamentos de células (filamentoso) se alimenta por fototrofia ou osmotrofia da mesma maneira como uma célula única. Mas imaginem uma ameba que se mova e se alimente (por fagocitose) através da emissão de pseudópodes, ou melhor, por que não ver a cena? É evidente que uma ameba não pode se unir a outra ameba e manter o mesmo modo de locomover-se e alimentar-se. E o mesmo ocorre em muitos outros protozoários e, de maneira geral, nos organismos fagotróficos, que obtêm os nutrientes por ingestão de organismos inteiros ou partes sólidas de organismos. Neste caso, tornar-se multicelular é verdadeiramente um problema! Não espanta que amebas e outros protistas fagótroficos que exibem multicelularidade formem corpos de frutificação, que produzem e dispersam esporos, mas não se alimentam. Desse modo, o conflito entre a necessidade de alimentar-se e a agregação celular é eliminado.
Contudo, para os animais, que são fagotróficos e descendem de linhagens fagotróficas, esta não poderia ser uma saída evolutiva: eles devem alimentar-se na condição multicelular. Aí está o grande quebra-cabeça da evolução animal, posto em destaque por Cavalier-Smith ao relembrar-nos de alguns conhecimentos básicos de biologia, que por vezes ficam perdidos num mundo de detalhes (uma questão curricular que é bom assunto para outra postagem). Qual vantagem seletiva poderia contrabalançar a perda de eficiência nutricional que decorre de células fagotróficas se unirem umas às outras? Uma perda de aptidão evolutiva (fitness) pode ser vislumbrada neste caso; logo, qual ganho de fitness poderia compensá-la?
Assim, a multicelularidade animal é única, no sentido de que coloca um problema evolutivo muito específico. Isso ajuda a explicar por que, a despeito da facilidade com a qual bactérias e protistas se tornam multicelulares, os animais evoluíram uma vez apenas na história da vida. Para um fagótrofo tornar-se multicelular, ele deve passar da fagocitose celular, que lhe permite, na condição unicelular, englobar outros organismos ou pedaços deles, para a alimentação por meio de uma boca e um sistema digestivo multicelulares. É fácil ver que isso não é trivial. Trata-se da formação de uma estrutura formada por várias células numa geometria específica, com novas funções e estruturas celulares, ausentes nos ancestrais unicelulares. Estes novos tipos celulares devem cooperar num nível organizacional superior, a maioria deles abandonando a capacidade de alimentar-se e reproduzir-se, desvantagens seletivas ao nível da célula individual que devem ser compensadas por vantagens seletivas ao nível do organismo multicelular como um todo.
A lição importante NÃO é que há aí alguma complexidade irredutível que o darwinismo ou mesmo o evolucionismo não pode explicar, como o cantochão do design inteligente gosta de insistir. A lição não é a aceitação de um planejador inteligente que, em termos naturalistas, nada explica (ou seja, se o planejador for alguma divindade, esta ideia, que pode ser válida da perspectiva de certas visões de mundo, não é científica, e não tem lugar no ensino de ciências, contrariamente ao que pensam os defensores do design inteligente, ou se for alguma entidade natural, digamos, algum tipo de ET, temos aí uma redução ao absurdo do design inteligente, porque implicaria uma sucessão ao infinito de ETs – o peso de ter de propor uma explicação está nos ombros dos defensores do design inteligente e nenhuma boa explicação até hoje saiu dessa seara). Já do lado do pensamento evolutivo, temos boas explicações. A lição, pois, é que precisamos conhecer mais sobre evolução para entender como os animais evoluíram a partir de ancestrais fagotróficos.
Muitos aspectos interessantes da explicação evolutiva aparecem neste caso. Primeiro, uma transição evolutiva fundamental na história da vida, como a da unicelularidade para a multicelularidade, implica conflito (trade-off) entre seleção ao nível do indivíduo unicelular anterior e seleção ao nível do indivíduo multicelular em formação. Como se resolve esse conflito? Trata-se de comparar a força da seleção nos dois níveis de individualidade. Se as vantagens dos unicelulares forem maiores, a multicelularidade não se torna uma solução evolutiva estável, a não ser que algum modo de acomodar suas desvantagens surja (como no caso da multicelularidade reprodutiva, mas não trófica, das amebas e de outros protistas, que mencionamos acima). Se a multicelularidade se tornou uma solução evolutiva estável, então as vantagens seletivas do novo individuo multicelular foram maiores do que as desvantagens dos indivíduos unicelulares que passaram da condição de vida livre à condição de células contidas num indivíduo situado num nível de organização superior.
Uma segunda lição deste caso é óbvia: é claro que, em certo momento da história evolutiva, os indivíduos multicelulares tiveram maiores vantagens seletivas do que os unicelulares. Afinal, existem multicelulares! Este fato óbvio coloca, então, o quebra-cabeças de explicar como isso ocorreu no caso dos animais: como se formou um animal multicelular com vantagens seletivas que superaram as desvantagens dos unicelulares ancestrais?
Daí decorrem novas lições fundamentais para quem quer entender evolução. É preciso situar a linhagem de multicelulares dos quais estamos falando na árvore da vida, para entender de qual grupo de organismos eles evoluíram, ou, de modo mais correto, qual seu grupo-irmão, com o qual compartilham um ancestral comum. É preciso encontrar características que estabelecem uma continuidade evolutiva entre a condição unicelular e a multicelular, características compartilhadas que permitam explicar como houve a transição de uma condição a outra. Aqui mais conhecimento é necessário: estas características compartilhadas não são geralmente óbvias, não são a mesma coisa – bioquímica, morfológica, funcionalmente – numa e noutra linhagem, mas frequentemente são moléculas ou estruturas compartilhadas que, numa linhagem, cumpre certa função, tipicamente com algum papel em sua evolução, e na outra, foram cooptadas, foram aproveitadas para outra função.
A colocação dos animais na árvore da vida é, portanto, um requisito fundamental para entender de onde eles vieram (e, claro, de onde nós viemos). Em 1866, Henry James-Clark descobriu os coanoflagelados e seu modo de alimentar-se de bactérias aprisionadas por um colar (formado por microvilosidades recobertas por muco) que cerca um flagelo ondulante, o qual gera uma corrente de água que leva as bactérias na direção do corpo celular, que então as fagocita (Figura 1). Ele notou que as esponjas possuem células com colares similares, com a mesma estrutura e o mesmo modo de alimentação, os coanócitos. Isso o levou a sugerir que esponjas haviam evoluído a partir de coanoflagelados. Esta ideia passou por ondas de aceitação e rejeição na história da biologia. Quando fiz graduação em biologia, era uma ideia rejeitada, uma especulação baseada numa similaridade morfológica enganadora. Não obstante, a hipótese de James-Clark tem sido apoiada pelas melhores árvores evolutivas hoje disponíveis, baseadas em muitas sequências gênicas ou proteicas. Estas árvores indicam que (1) as esponjas são aparentadas dos outros animais (também uma ideia que já foi questionada) e (2) os coanoflagelados são os parentes mais próximos dos animais (a título de exemplo, os leitores podem ver o artigo Monophyletic origins of metazoa [Origens monofiléticas dos metazoários], no qual também terão conhecimento de que os fungos são, entre os multicelulares, o grupo mais aparentado dos animais).

Se considerarmos os animais hoje existentes, as dificuldades resultantes da alimentação de um organismo fagotrófico multicelular só poderiam ser superadas pelas esponjas. Estes animais poderiam evoluir de protistas sem mudar seu modo de alimentação. O problema da origem dos animais pode ser traduzido, então, no problema de como e por que as esponjas evoluíram de coanoflagelados e deram posteriormente origem a todos os outros animais.
O primeiro passo para resolver este problema é considerar a transição coanoflagelado/esponja. O foco deve recair, naturalmente, sobre coanoflagelados coloniais, dos quais provavelmente multicelulares evoluíram. Em colônias de coanoflagelados, todas as células podem alimentar-se. O tipo de colônia de coanoflagelados do qual as esponjas mais provavelmente evoluíram são esferas celulares planctônicas (Figura 1). A transição para a forma do corpo de uma esponja implica que parte das células devem abandonar a alimentação por meio do colar, tornando-se células de fixação ao substrato, dando origem, assim, aos pinacócitos das esponjas. Isso diminui a eficiência de alimentação da nova forma multicelular. O que compensaria esta perda de fitness? Esta, como vimos acima, é uma pergunta fundamental.
Como argumenta Cavalier-Smith, uma esponja somente poderia evoluir se possuísse um corpo que mais do que compensasse, por um aumento indireto na eficiência de sobrevivência e reprodução, a perda direta na eficiência da alimentação, em vista da menor proporção de células com função trófica. Trata-se de definir as forças seletivas que favoreceram um organismo que exibia uma diferenciação entre coanócitos, com função trófica, e células sem esta função, como os pinacócitos das esponjas atuais. A hipótese é que tais forças seletivas favoreceram formas corporais sésseis capazes de filtrar maiores volumes de água, compensando a perda na eficiência alimentar decorrente da diferenciação de um tipo celular sem função trófica.
Podemos chegar, desse modo, ao modelo mostrado na Figura 1, de acordo com o qual esponjas evoluíram a partir de esferas multicelulares planctônicas, encontradas em coanoflagelados da ordem Craspedida, que apresentam uma superfície formada por uma monocamada de células unidas por seus corpos celulares, capazes de se alimentar do mesmo modo que coanoflagelados unicelulares. Este modelo evolutivo é, claro, de natureza hipotética e é preciso examinar o peso da evidência a seu favor. De qualquer modo, temos aí um modelo plausível para a transição coanoflagelado-esponja, na diferenciação de um tipo celular somático capaz de se ancorar num substrato rochoso e de secretar matriz extracelular – que corresponderia, nas esponjas atuais, a um pinacócito basal. A matriz formada por mucopolissacarídeos e colágeno formaria um mesoílo, que ajudaria a suportar a estrutura multicelular, capaz de filtrar maiores volumes de água, obtendo mais alimento e, assim, superando competitivamente simples colônias ramificadas de coanoflagelados. Desse modo, os custos evolutivos da perda da capacidade de filtração pelos pinacócitos basais e da secreção da matriz extracelular poderiam ser compensados. Com a continuidade da pressão seletiva por capacidade de filtração, os demais passos evolutivos mostrados na Figura 1 poderiam ter lugar, levando a corpos multicelulares ainda maiores e à oogamia.
As pré-esponjas teriam se diversificado, segundo Cavalier-Smith, na Biota Ediacarana (também conhecida como Biota Vendiana), formada por fósseis com aspecto tubular e sésseis em forma de frondes que viveram entre ca. 635 e 542 milhões de anos atrás, antes da Explosão do Cambriano, quando teve origem a maioria dos filos animais. A partir delas, teriam evoluído as esponjas, dotadas de um sistema aquífero formado por óstios, poros no corpo, por onde entra a água, e ósculos, por onde sai a água filtrada pelos coanócitos nas cavidades do corpo, a espongiocele, para obtenção de alimento. Cavalier-Smith discute em seu artigo uma hipótese para a origem das esponjas a partir das pré-esponjas por ele postuladas, assim como a origem de cnidários, ctenóforos (em outras hipóteses sobre a evolução dos animais, colocados como grupo mais basal do que as esponjas) e placozoários (também considerados animais basais em outras hipóteses). Ele se debruça também sobre a evolução do celoma, que considera um caráter primitivo dos animais, e não derivado, de modo similar à ideia de que organismos triblásticos seriam mais primitivos do que as linhagens diblásticas. Ele discute a origem dos animais bilaterais, do sistema nervoso, de diferentes classes de cnidários. Em suma, uma grande diversidade de assuntos é abordada em seu recente artigo, sobre os quais não temos como nos debruçar aqui.
Não há dúvida de que os modelos propostos por Cavalier-Smith suscitam uma série de controvérsias. Mas é exatamente porque têm qualidade e consistência como proposições científicas que merecem discussão. Similarmente, ele se contrapõe a outras hipóteses propostas na literatura científica, questionando-as exatamente porque merecem questionamento. Não podemos certamente acusar Cavalier-Smith de não exibir uma capacidade de síntese imaginativa, a qual é fundamental para que tenhamos hipóteses científicas consistentes sobre as quais nos debruçarmos. Estas hipóteses são infinitamente melhores do que outros exercícios mais vãos que se tornaram populares nas últimas décadas.
Charbel N. El-Hani
(Instituto de Biologia/UFBA)
PARA SABER MAIS:
Cavalier-Smith, T. (2010). Deep phylogeny, ancestral groups and the four ages of life. Philosophical Transactions of the Royal Society B 365: 111-132.
Cavalier-Smith, T. (2017). Origin of animal multicellularity: precursors, causes, consequences—the choanoflagellate/sponge transition, neurogenesis and the Cambrian explosion. Philosophical Transactions of the Royal Society B 372: 20150476.
Dayel, M. J., Alegado, R. A., Fairclough, S. R., Levin, T. C., Nichols, S. A., McDonald, K. & King, N. (2011). Cell differentiation and morphogenesis in the colony-forming choanoflagellate Salpingoeca rosetta. Developmental Biology 357: 73–82.
Shen, S., Zhu, M., Wang, X., Li, G., Cao, C. & Zhang, H. (2010). A comparison of the biological, geological events and environmental backgrounds between the Neoproterozoic-Cambrian and Permian-Triassic transitions. Science China Earth Sciences 53: 1873-1884.
Wainright, P. O., Hinkle, G., Sogin, M. L. & Stickel, S. K. (1993). Monophyletic origins of the Metazoa: An evolutionary link with Fungi. Science 260: 340-342.
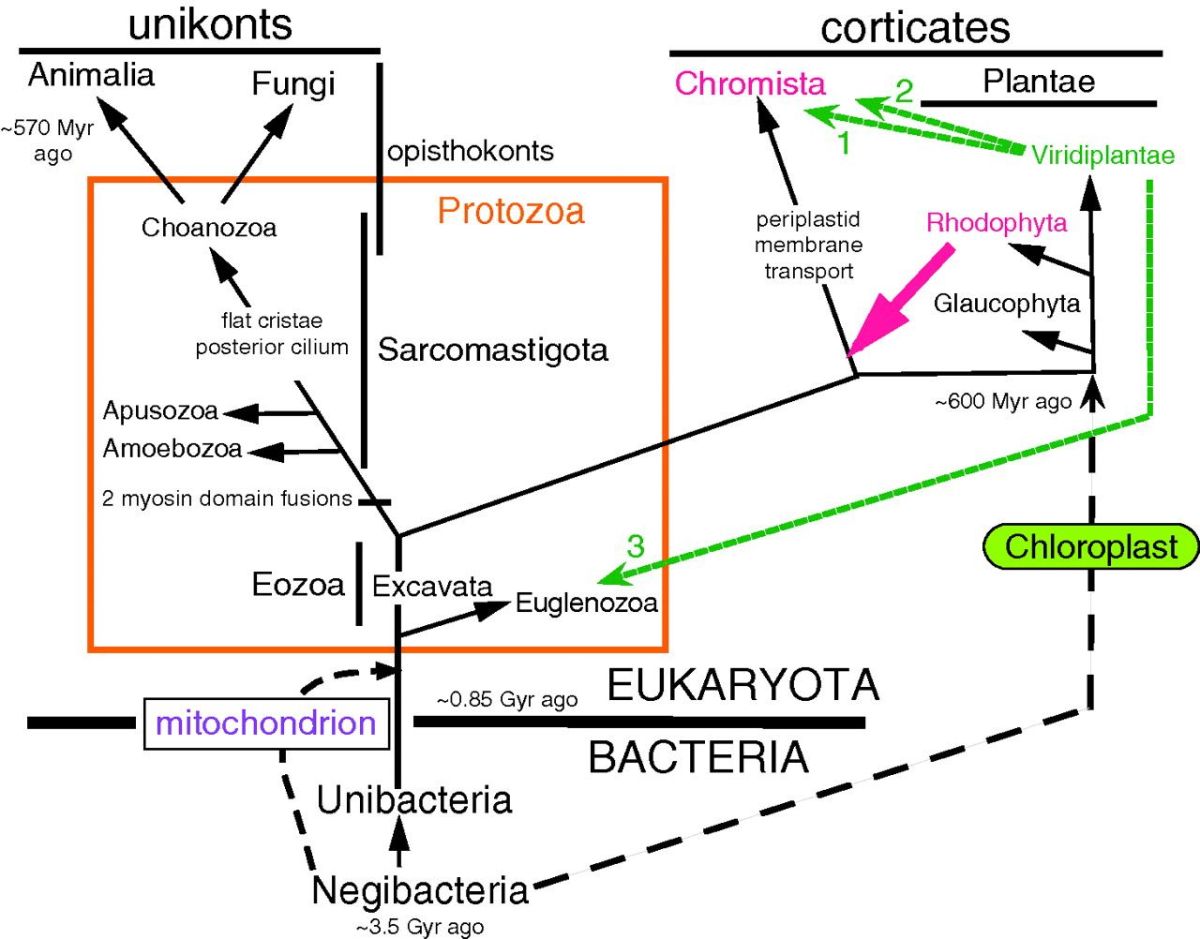
Figura na abertura: Árvore evolutiva dos eucariotos. Os Reinos Plantae (as plantas verdes) e o supergrupo Chromista (que inclui todas as algas cujos cloroplastos contêm clorofilas a e c, assim com todas as formas sem cloroplastos intimamente aparentadas) constituem uma primeira irradiação de eucariotos, originados independentemente dos Reinos Animalia (os animais) e Fungi (os fungos), que correspondem a outra irradiação de eucariotos. Estes últimos tiveram origem a partir de Choanozoa (os chamados coanoflagelados) (de Cavalier-Smith, 2009).

Charbel, belo texto e uma bela síntese de um artigo interessante. Eu preciso ler o trabalho em questão, mas acredito que de fato ele deve contribuir para novas ideias acerca da evolução dos animais. Como você cita em seu trabalho, a origem dos animais já foi alegada para vários grupos de eucariotos unicelulares (antes conhecidos como protistas – termo este que talvez devesse ser abandonado, uma vez que remete a um “reino” que hoje sabemos não existir). Coanoflagelados têm sido os principais suspeitos, pois a similaridade morfológica (colarinhos e flagelo) com os coanócitos das esponjas indicariam que essa transição, como proposta por Cavalier-Smith (mas já sugerida algumas vezes no passado, por ex. por Nielsen em 2008 – Evol. Dev. 10: 241-257) parece ser natural. Eu tenho alguns “pés atrás” para estas ideias evolutivas gradistas. A começar pela “árvore” que é apresentada no começo do texto, no qual linhagens que tiveram o mesmo tempo de evolução e estão atualmente vivas, aparecerem como “degraus” de uma escada evolutiva, no qual uma linhagem parece ter dado origem a outra. Além disso, aparentemente a morfologia das células (coanócito e coanoflagelado) parece ser uma adaptação estrutural para exercer a função que elas de fato exercem (criar uma corrente de água para trazer o alimento para a célula, veja Mah et al. 2014. Evol. Dev. 16:25-37). Juntando ao fato de que a presença das esponjas como sendo a linhagem viva que primeiro divergiu dos demais metazoários tem sido questionada em trabalhos com muita consistência, este argumento de Cavalier-Smith precisa ser estudado e absorvido com muita cautela. Enfim, eu não li o artigo e estou comentado o seu texto baseado no que conheço sobre as esponjas. Vou ler com cuidado e carinho o trabalho tema desta postagem e posso criar um novo texto para continuarmos com este debate. De qualquer forma, concordo com você: é preciso sairmos do círculo vicioso de não propormos coisas novas. É preciso que estas novas proposições sejam feitas com bons argumentos que são construídos a partir de evidências científicas e que possa, como é o cerne da ciência, serem testados.
Abraços,
Emilio
CurtirCurtir
Aproveitando o “gancho ” do artigo: li um artigo sobre simulações de possíveis “animais extraterrestres ” de pesquisadores do Reino Unido, conhecidos como os Aliens de Darwin e para suas simulações os pesquisadores precisavam pressupor transições como essas postuladas pelo senhor e o autor acima e a seleção natural é claro. É incrível a diversidade de morfologia possível de hipotéticos animais em outros mundos simulados. Se o senhor pudesse tecer alguns comentários sobre essas simulações ficaria muito grato. Se o senhor puder é claro. Abc
CurtirCurtir
Como professor de biologia vejo muito os alunos imprecionarem com a possibilidade de vida extraterrestre. Muitas vezes não tenho fundamentos para uma discussão interessante sobre o assunto. Um aluno um dia desses me perguntou se as simulações dos Aliens de Darwin tinham condições de existência em outros planetas que não a terra. Disse que eram simulações e modelos de formas de vida possíveis com certas premissas ambientais de antemão é claro e com base nas formas de vida baseada no carbono e em vida multicelulares… Se o senhor puder tecer alguns comentários.
CurtirCurtir
Os estudos de vida artificial são bem importantes, Dalmir, para investigarmos não somente a vida tal como a conhecemos na Terra, mas também a vida em todas as formas que ela poderia ter. As questões filosóficas levantadas também são muito interessantes: por exemplo, para os estudos de vida artificial, a vida é definida como forma, como organização, e não como uma base material específica. Um exemplo: material genético não precisa ser constituído por ácido nucléico na vida tal como ela poderia ser. Estes estudos têm sim relevância para pensarmos a pesquisa sobre vida extraterrestre. Agora, note, é importante esclarecer para os alunos que, em campos científicos como a astrobiologia, a busca não é por ETs, vida inteligente etc. Conforme a equação de Drake (https://en.wikipedia.org/wiki/Drake_equation), a probabilidade de haver vida em outros planetas é muito alta, mesmo a probabilidade de haver vida inteligente é muito alta. Pense na vastidão do universo. Se você delimitar um pequeno quadrado no céu e olhar para ele você estará vendo milhares de estrelas. Agora multiplique isso pela quantidade de pequenos quadrados que você pode situar no céu. São tantos tantos tantos mundos possiveis que, sim, é plausível que não estejamos sozinhos no Universo. Mas note que isso não significa que ETs estejam nos visitando, ou que vida inteligente será achada facilmente, ou mesmo que vida seja achada facilmente. Pense no tamanho do desafio para nós: a vastidão é inominável, as distâncias são intransponíveis. Para chegar mesmo em locais não muito distantes do universo, com a tecnologia que temos hoje, morreríamos no caminho porque o tempo seria maior do que o tempo da vida humana. Agora imagine as mesmas dificuldades para possíveis ETs. Não podemos dizer que seria impossível chegarem aqui, caso existam, mas não podemos também achar que é um almoço de graça, a piece of cake, cruzar a vastidão do espaço para fazer contato. Fundamental distinguir para seus alunos o que é o estudo científico da vida extraterrestre (como na exobiologia e na astrobiologia), e o que é ficção científica e wishful thinking.
Abraços
Charbel
CurtirCurtir
Obrigado professor. Já tenho agora uma direção quando alunos me perguntarem se Etes vivem no meio de nós. Abçs.
CurtirCurtir
Gostei muito da sua resposta e adorei o blog. Sou professora de biologia evolutiva numa universidade em Moçambique. Parabéns pela simplicidade dos artigos mantendo a qualidade cientifica dos mesmos.
CurtirCurtir
Excelente, Cristina! Ficamos muito felizes do impacto e de sabermos que estamos alcançando nossos objetivos no blog.
CurtirCurtir
À respeito do surgimento dos organismos multicelulares, eu tenho algumas dúvidas: como os organismos unicelulares individuais que constituem uma colônia de células e que possuem material genético diferente uns dos outros passam a apresentar o mesmo genoma e, por consequência, se tornam células de um mesmo organismo multicelular individual? Esse processo ocorre por meio da transmissão horizontal de genes, assim como nas bactérias? Por fim, as células do novo organismo multicelular passam a apresentar o mesmo antes ou depois da diferenciação e especialização das células em funções diferentes? No demais, parabéns pelo artigo de excelente qualidade. Eu o agradeço por ter satisfazido a minha curiosidade sobre este assunto fascinante.
CurtirCurtir
Olá, tipicamente organismos multicelulares se formam a partir de colônias de indivíduos da mesma espécie, de modo que essa conversão para o mesmo genoma não é um passo necessário. Sendo indivíduos de espécies diferentes, podem formar outros tipos de associações, mas não alçam à condição de organismo multicelular, até onde sei.
Abs
Charbel
CurtirCurtir
Muito obrigado pela resposta. Havia até me esquecido que tinha postado este comentário, especialmente depois que eu tive que apagar o gmail/e-mail que eu usei para postá-lo por conta de um bug no Google, por isso tive que criar outro. Quanto à sua resposta, ela acabou me deixando com uma outra dúvida: como então os atuais animais — pelo menos em seus primeiros estágios de desenvolvimento — possuem células que possuem o mesmo material genético?
CurtirCurtir
Olá, Observador da Via Láctea
O material genético é compartilhado através de espécies, clados, domínios, em formas homólogas, porque descendentes das mesmas sequências ancestrais. Através dos padrões de herança e especiação, sequências descendentes estabelecem uma árvore de genes que não está restrita a barreiras entre domínios, espécies ou clados.
Abs
Charbel
CurtirCurtir
Ótimo artigo. Belo trabalho professor Charbel!!!
CurtirCurtir