
Desde a educação básica, aprendemos como distinguir procariontes e eucariontes. Procariontes incluem bactérias e arqueas, organismos cujo genoma se resume a um único cromossomo circular, que não se encontra num núcleo separado do citoplasma. Eucariontes têm mais de um cromossomo linear, dentro de um núcleo separado do citoplasma por uma membrana dupla. Procariontes têm ribossomos e citoesqueleto, que são distintos daqueles dos eucariontes, e não possuem organelas com membrana.
Mitocôndrias e cloroplastos estão entre as organelas mais conhecidas dos eucariontes. Mitocôndrias são encontradas em todos os eucariontes… Ou ao menos era o que achávamos até ter sido publicado, pouco mais de um mês atrás, o primeiro relato de um eucarioto sem mitocôndria, Monocercomonoides sp. (uma nova espécie, ainda sem nome específico). No momento, é uma exceção, dessas tão comuns na biologia. Nada há de espantoso em tais exceções: generalizações biológicas, como a de que todos os eucariotos têm mitocôndrias, são produtos da evolução. E a própria evolução gera novidades que limitam a validade dessas generalizações. É o caso de Monocercomonoides sp.: ele perdeu secundariamente mitocôndrias que haviam sido adquiridas por seus ancestrais e, com isso, se tornou uma exceção na generalização de que todos os eucariotos possuem mitocôndrias.
A evolução de mitocôndrias é entendida por meio de uma teoria presente no conhecimento escolar: a teoria endossimbiótica serial. Essa teoria afirma que mitocôndrias, responsáveis pela respiração celular, e cloroplastos, responsáveis pela fotossíntese, se originaram a partir de bactérias simbiontes.
Vale esclarecer alguns termos ecológicos antes de prosseguirmos. O termo “simbiose” designa uma relação ecológica na qual duas espécies vivem associadas por longos períodos. Numa relação simbiótica, pelo menos uma espécie se beneficia, enquanto a outra pode ser prejudicada (falamos então em parasitismo), não ser afetada (comensalismo) ou ser beneficiada (mutualismo). Quando uma relação simbiótica mutualista ocorre entre um hospedeiro e um organismo que vive em seu interior, falamos em endossimbiose.
A teoria endossimbiótica serial propõe que a evolução da célula eucariótica envolveu a incorporação de vários endossimbiontes, que perderam sua autonomia e se tornaram organelas. Temos aí uma novidade no pensamento evolutivo, que enfatizou desde Darwin a divergência de descendentes a partir de ancestrais comuns, expressa de modo sugestivo na metáfora de uma árvore da vida com diversos ramos. A teoria endossimbiótica enfatiza a fusão de ramos na árvore da vida, dando origem a novas espécies, no fenômeno da simbiogênese. Esta não é uma ideia nova.
O botânico russo Konstantin Mereschkowskyi (1855-1921) foi o primeiro a propô-la, ao defender num artigo de 1905 a origem endossimbiótica do cloroplasto e do núcleo.
Em 1918, o cientista francês Paul Portier (1866-1962) publicou Les Symbiotes (Os simbiontes), no qual propunha que mitocôndrias tinham origem simbiótica.
O biólogo norte-americano Ivan Wallin (1883-1969) também estendeu a ideia a mitocôndrias no livro Symbionticism and the origin of species (Simbionticismo e a origem das espécies), de 1927.
A conexão com a evolução darwiniana foi destacada pelo botânico russo Boris Mikhailovich Kozo-Polianski (1890-1957) em seu livro Simbiogênese: um novo princípio da evolução. A simbiogênese se configuraria como um novo princípio por basear a seleção natural no fenômeno da simbiose.
Todos esses desenvolvimentos exerceram pouca influência sobre o pensamento evolutivo que se consolidou dos anos 1930 a 1950 na teoria sintética da evolução, que não atribuiu qualquer papel à simbiogênese. Coube à bióloga norte-americana Lynn Margulis (1938-2011) retomar a ideia da simbiogênese, tornando-a muito mais influente do que jamais fora. Em parte, isso resultou de uma série de evidências empíricas favoráveis à simbiogênese que Margulis reuniu a partir dos anos 1960. Por exemplo, que mitocôndrias e cloroplastos têm DNA próprio, que se replica independentemente do DNA nuclear. Que essas organelas são capazes de reprodução no interior da célula. Que possuem ribossomos similares aos de procariontes, entre outras evidências. Assim, a ideia da origem evolutiva de mitocôndrias e cloroplastos por endossimbiose foi ganhando aceitação crescente, apesar das controvérsias. Hoje, ela é largamente aceita, como mostra sua presença no conhecimento científico escolar como ideia estabelecida pela ciência.
Não foi um mar de rosas para Margulis. Como comenta Richard Dawkins, sua coragem na defesa da teoria endossimbiótica foi admirável. Ela a levou de uma ideia não-ortodoxa à ortodoxia do pensamento evolutivo. Em 1966, Margulis escreveu On the origin of mitosing cells (Sobre a origem das células mitóticas), ainda assinando Lynn Sagan (Ela foi casada com Carl Sagan), um artigo rejeitado por 15 periódicos científicos até ser publicado pelo Journal of Theoretical Biology, para tornar-se um marco na história da teoria endossimbiótica. Em 1970, ela desenvolveu mais a teoria em Origin of Eukaryotic Cells (Origem das células eucarióticas). As cinco décadas seguintes veriam Margulis envolvida em outros desenvolvimentos científicos importantes, alguns deles também controversos, como a teoria Gaia e a proposta da divisão da vida em cinco Reinos.
Considerando todo seu conteúdo, a teoria endossimbiótica não se tornou de fato uma ortodoxia. Apenas a explicação endossimbiótica da origem de mitocôndrias e cloroplastos foi aceita. Mas a teoria que remonta aos trabalhos originais de Mereschkowskyi e Wallin, que foi defendida arduamente por Margulis, propõe que toda a célula eucariótica se originou da fusão de células procarióticas ancestrais. Não seriam somente mitocôndrias que teriam se originado de alfaproteobactérias simbiontes (de qual grupo de alfaproteobactérias exatamente ainda é controverso, como discutido num artigo no periódico Scientific Reports, e num artigo do qual sou um dos autores, em PLOS One). Não seriam somente os cloroplastos que tiveram múltiplas origens, num evento primário de endossimbiose com uma cianobactéria, e em diversos eventos secundários e terciários. O núcleo teria se originado de uma arquea simbionte. Os flagelos, de espiroquetas, e assim por diante. A célula eucariótica seria uma comunidade ecológica que se tornou um organismo individual.
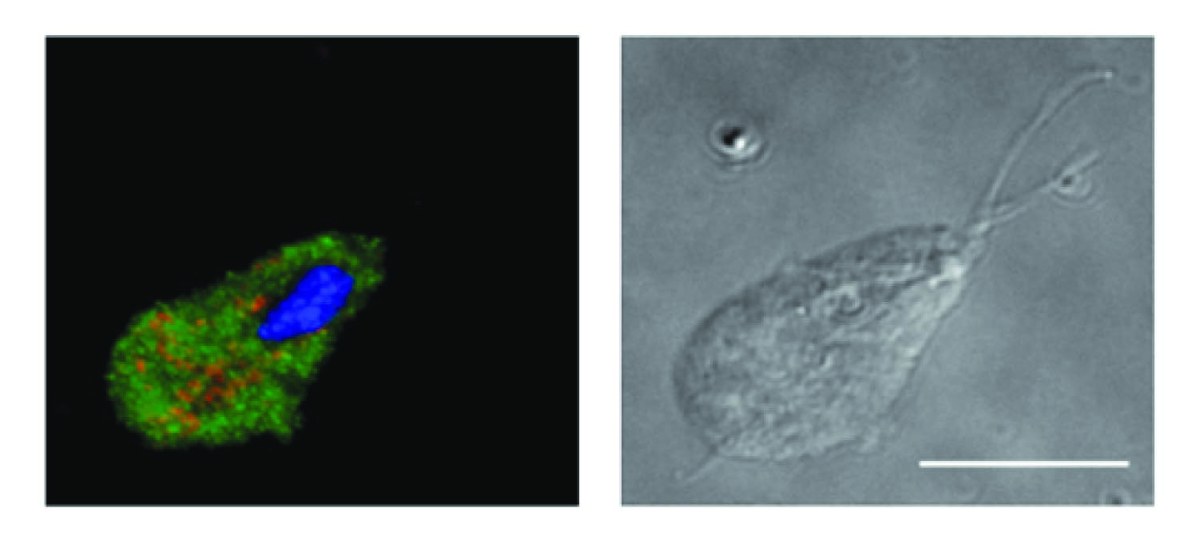
Seria a importância evolutiva da simbiogênese afetada pelo achado de um eucarionte sem mitocôndrias? Não é o caso. O principal impacto do artigo A eukaryote without a mitochondrial organelle, de Anna Karnkowska e colaboradores, é que ele mostra que mitocôndrias não são absolutamente essenciais para a viabilidade da célula eucariótica. Mas como o microorganismo funciona sem mitocôndrias? Como o metabolismo energético ocorre sem mitocôndrias em Monocercomonoides sp.? Como também ocorre em organismos com mitocôndrias muito reduzidas, o genoma dessa espécie não codifica enzimas envolvidas no metabolismo energético aeróbio, por exemplo, do ciclo de Krebs ou da cadeia respiratória. Trata-se de metabolismo energético bem diferente daquele que aprendemos nos livros didáticos. É através de uma glicólise estendida que Monocercomonoides sp. produz ATP, a moeda energética da célula. É idêntico ao que ocorre em espécies como Giardia intestinalis e Entamoeba histolytica, que têm mitocôndrias bastante reduzidas e produzem ATP apenas no citoplasma.
A história da ciência nos aconselha, contudo, a ter cautela diante da descoberta. Nos anos 1980, foi proposto, na hipótese do Reino Archezoa, de Thomas Cavalier-Smith, que haveria microorganismos eucariotos que jamais teriam adquirido mitocôndrias e outras organelas, como peroxissomos. Na década seguinte, foi mostrado que havia mitocôndrias nas espécies do Reino proposto, mas elas haviam sido tão modificadas que se torna difícil reconhecê-las. A presença de mitocôndrias passou a ser verificada então por meio de proteínas marcadoras mitocondriais. Assim, a ausência de genes que codificam proteínas mitocondriais é a evidência mais importante em apoio à hipótese de que Monocercomonoides sp. é um eucarioto que não possui tais organelas. Os dados genômicos relatados no artigo indicam a ausência de DNA mitocondrial ou de genes mitocondriais translocados para o genoma nuclear nessa nova espécie.
Num bom trabalho científico, explicações alternativas devem ser consideradas. Karnkowska e colegas examinam a possibilidade de que exista em Monocercomonoides sp. uma mitocôndria com composição proteica totalmente alterada. Isso é possível, em princípio. Mas é uma hipótese que não leva a avanços, porque não teríamos maneira de reconhecer tal organela hipotética. Sem evidência positiva a favor dessa alternativa, podemos concordar com os autores: a completa ausência de marcadores e vias metabólicas mitocondriais é evidência legítima da ausência da organela.
É sempre interessante ver o que sabemos mudando bem debaixo de nossos narizes, mostrando o quanto ainda temos para conhecer acerca da vida.
Charbel N. El-Hani
(Instituto de Biologia/UFBA)
PARA SABER MAIS:
Karnkowska, A. et al. (2016). A Eukaryote without a Mitochondrial Organelle. Current Biology 26: 1-11.
Kozo-Polianski, B. M. (2010). Symbiogenesis: A New Principle of Evolution. Cambridge, MA: Harvard University Press.
Margulis, L. (1971). Origin of Eukaryotic Cells. New Haven, CT: Yale University Press.
Margulis, L. (2003). Acquiring Genomes: A Theory of the Origin of Species. New York, NY: Basic Books.
Martin, L. & Kowallik, K. (1999). Annotated English translation of Mereschkowsky’s 1905 paper ‘Über Natur und Ursprung der Chromatophoren imPflanzenreiche’. European Journal of Phycology 34: 287-295.
Sapp, J. (1994). Evolution by Association: A History of Symbiosis. Oxford: Oxford University Press.
Wallin, I. (1927). Symbionticism and the Origin of Species. Baltimore, MD: Williams & Wilkins Co.

“A célula eucariótica seria uma comunidade ecológica que se tornou um organismo individual”. Ótima frase, nunca tinha parado pra pensar e ver a célula assim. Só não achei que ficou claro como funciona essa “glicólise estendida”. Mas o texto é bem esclarecedor. Abraços.
CurtirCurtir
Oi Vinícius,
Obrigado pelo comentário e desculpe pela demora em responder. Ficamos contente que tenha apreciado o texto. O comentário nos dá oportunidade de incluir uma discussão que estava na primeira versão da postagem, mas tiramos por questão de espaço: Um tema evolutivo importante reverbera na ideia de que a célula eucariótica seria uma comunidade ecológica que se tornou um organismo individual: na evolução das espécies, novos níveis de individualidade emergem em transições evolutivas em que surgem novas totalidades, compostas por partes que haviam evoluído anteriormente. Esta é uma das razões para o que John Maynard Smith (1920-2004) e Eörs Szathmáry (n. 1959) denominam grandes transições evolutivas. Para que tal transição evolutiva ocorra, o balanço de forças entre a seleção ao nível dos componentes e do todo que eles estão formando deve se reconfigurar: a vantagem seletiva de um componente na competição com outros componentes deve ser menor do que a vantagem seletiva da totalidade que eles formam em competição com outras totalidades similares. Dito de modo sintético, a seleção ao nível da totalidade deve ser mais intensa do que a seleção ao nível dos componentes. Somente assim a cooperação entre as partes pode sobrepujar a competição entre elas, estabilizando o novo nível de individualidade que está se formando.
Quanto à glicólise estendida, esperamos fazer em breve uma postagem a respeito. Mas aqui está um aperitivo. Na glicólise, ATP é produzido, com rendimento menor do que na respiração celular, e cofatores como NAD são reduzidos, recebendo hidrogênio e elétrons de metabólitos intermediários da glicólise. Contudo, com o tempo a célula começa a não ter mais cofatores como NAD disponíveis e a glicólise será interrompida. Por isso, os cofatores devem ser reoxidados, tornando-se disponíveis novamente para o transporte de prótons (H+) e elétrons. Quando um eucarioto tem mitocôndrias, isso acontece no interior destas, na cadeia respiratória, quando os cofatores transferem os elétrons para a cadeia de citocromos e os prótons são bombeados a favor do gradiente de concentração para o espaço entre as duas membranas das mitocôndrias. Gera-se assim o gradiente que é aproveitado na síntese de ATP. Monocercomonoides não tem mitocôndrias, no entanto, e então necessita de passos metabólicos adicionais para reoxidar os cofatores, como ocorre em bactérias que fazem fermentação após glicólise, com essa mesma função. No caso de Monocercomonoides, os cofatores são reoxidados por fermentação que produz acetato e etanol, ou por meio da hidrogenase-[FeFe], produzindo gás hidrogênio. Pronto, isso é a glicólise estendida. Evitamos comentar na postagem porque envolve uma explicação bioquímica mais complicada, como deve ter ficado claro acima.
Um último comentário. A situação em Monocercomonoides é idêntica àquela em espécies mais conhecidas das pessoas, como Giardia intestinalis e Entamoeba histolytica, que são relativamente próximas, em termos evolutivos, de Monocercomonoides. A razão é similar: G. intestinalis e E. histolytica não perderam suas mitocôndrias, mas têm mitocôndrias muito reduzidas, que evoluíram independentemente, formando os chamados mitossomos, e também produzem ATP no citossol.
Abraços
Charbel
CurtirCurtir
Entendi que a ausência da mitocôndria não refuta a teoria, mas ficará claro para um estudante de ensino médio o que esta dito no título? Que o achado apóia a teoria? Não encontrei.
CurtirCurtir
Oi Débora,
Muito oportuna sua pergunta. De fato, nos dá a oportunidade de esclarecer o título que, admitimos, ficou um pouco críptico. A ideia foi a seguinte: o achado de um eucarioto sem mitocôndria poderia sugerir uma origem dos eucariotos que não envolveria endossimbiose, ao menos no caso dessa organela, mas, como a interpretação das evidências colhidas por Karnkowska e colaboradores aponta ter sido perda secundária de mitocôndrias numa linhagem cujos ancestrais possuíam mitocôndrias, o achado termina por reforçar a hipótese da evolução das mitocôndrias por endossimbiose, porque sua interpretação pressupõem tal hipótese, ao se referir a uma perda secundária.
Abraços
Charbel
CurtirCurtir
Bom dia Charbel
Parabenizo a você e toda equipe pela iniciativa do blog. Um bom canal de discussão e informações confiáveis pelo que conheço de você. Em relação a esta publicação não estaria se referindo a organismos que ao invés de ter mitocôndria tem hidrogenossomos? Infelizmente não consegui o artigo “A Eukaryote without a Mitochondrial Organelle” completo. Em caso afirmativo, está não seria uma descoberto tão recente. Segundo Nigel Yarlett e Johannes H. P. Hackstein em “Hydrogenosomes: One Organelle, Multiple Origins” (BioScience Vol. 55, No. 8, 2005) estas organelas foram descritas em Tritrichomonas foetus por Lindmark e Müller em 1973. Estão relacionadas com a produção de ATP em eucariotos que não tem mitocôndria. Ainda segundo estes autores, hidrogenossomos e mitocôndrias não coexistem. Há inclusive uma discussão se hidrogenossomos não derivam de mitocôndrias ou se evoluíram independentemente de ancestral comum.
Abraços
José Roberto
CurtirCurtir
Oi José Roberto,
Bom vê-lo por aqui. Siga nosso blog!
O caso estudado pelos autores do artigo é diferente. Não tem nem hidrogenossomos. Vale conferir o original.
Abs
Charbel
CurtirCurtir